
Centre d'expertise sur les mammifères marins : Rapport de recherche scientifique 2006-2008
Table des matières
- Remerciements
- Message du directeur du Centre d'expertise sur les mammifères marins
- 1.0 Aperçu
- 2.0 Dynamique des populations
- 2.1 Abondance
- 2.1.1 SPLASH: Étude internationale sur les rorquals à bosse dans le Pacifique Nord (John Ford)
- 2.1.2 Abondance du béluga de l'ouest de la baie d'Hudson (Pierre Richard)
- 2.1.3 Abondance du béluga de l'estuaire du Saint-Laurent (Jean-François Gosselin)
- 2.1.4 Structure par âge, croissance, mortalité et densité de la population de bélugas dans l'Arctique canadien (Steve Ferguson)
- 2.1.5 Réduction soutenue de la croissance démographique du phoque gris (Don Bowen et Mike Hammill)
- 2.1.6 Moment de la mise bas et abondance des phoques à capuchon de l'Atlantique Nord-Ouest (Garry Stenson et Mike Hammill)
- 2.1.7 Abondance de l'otarie de Steller en Colombie-Britannique (Peter Olesiuk)
- 2.1.8 Abondance des baleines boréales de l'est de l'Arctique (Larry Dueck)
- 2.2 Établissement de la structure de la population à l'aide de la génétique moléculaire
- 2.2.1 Différencier les bélugas de l'est de la baie d'Hudson de ceux de l'ouest de la baie d'Hudson au moyen de marqueurs moléculaires (Lianne Postma)
- 2.2.2 La génétique moléculaire pour l'identification des stocks de narvals au Nunavut (Lianne Postma)
- 2.2.3 Modes de différenciation génétique chez les baleines boréales de l'est de l'Arctique canadien et de l'ouest du Groenland (Lianne Postma)
- 2.3 Reproduction et mortalité
- 2.3.1 Tactique d'accouplement des phoques gris mâles selon leur condition: l'importance de la taille du corps (Don Bowen)
- 2.3.2 Évaluation du dénombrement de nouveau-nés d'otaries de Steller à partir de photographies 35 mm prises à angle oblique (Peter Olesiuk)
- 2.3.3 Cycle biologique et dynamique des populations d'épaulards résidents du nord de la Colombie-Britannique (Peter Olesiuk, Graeme Ellis et John Ford)
- 2.4 Rôle des maladies dans les populations de mammifères marins
- 2.1 Abondance
- 3.0 La place des mammifères marins dans l'écosystème
- 3.1 Répartition
- 3.2 Habitudes
- 3.2.1 Caractéristiques de plongée et estimations de l'observabilité des baleines boréales de l'est de l'Arctique d'après la télémesure par satellite (Larry Dueck)
- 3.2.2 Écologie du phoque annelé (Steve Ferguson)
- 3.2.3 Mise au point de la méthode de surveillance acoustique passive pour surveiller de manière non intrusive les baleines dans leur environnement avec des exemples provenant de la voie navigable du Saint-Laurent et de l'Arctique (Yvan Simard)
- 3.3 Écologie alimentaire et régime alimentaire
- 3.4 Interactions entre le prédateur et la proie
- 3.5 Habitat des mammifères marins
- 3.5.1 Les phoques communs à Terre-Neuve et au Labrador: nouvelles données (Becky Sjare)
- 3.5.2 Caractéristiques sexospécifiques des troupeaux de loutres de mer de même sexe (Linda Nichol)
- 3.5.3 De la physique aux baleines: l'exemple de l'aire d'alimentation des cétacés à fanons de l'estuaire du Saint-Laurent (Yvan Simard)
- 4.0 Répercussions anthropiques sur les mammifères marins
- 4.1 Chasses commerciales et chasses de subsistance
- 4.2 Formulation d'avis scientifiques sur les TAC
- 4.2.1 Gestion du phoque du Groenland, du phoque à capuchon et du phoque gris au Canada (Garry Stenson)
- 4.2.2 Mortalité causée par la chasse chez les phoques à capuchon de l'Atlantique Nord-Ouest (Garry Stenson)
- 4.2.3 Détermination du total autorisé des captures de bélugas et de narvals du Nunavut (Pierre Richard)
- 4.3 Prises accessoires de mammifères marins
- 4.4 Relevés sismiques et sonars militaires
- 4.5 Les mammifères marins comme sentinelles de la contamination environnementale
- 4.6 Événements d'échouement et de mortalité
- 4.6.1 Mortalité causée par la chasse chez les phoques du Groenland dans l'Atlantique Nord-Ouest (Garry Stenson)
- 4.6.2 Bélugas de l'estuaire du Saint-Laurent – Examen du programme des carcasses (Lena Measures)
- 4.6.3 Réduction du risque de collision entre les navires et les baleines noires dans la région du bassin Roseway du sud-ouest de la plateforme Scotian (Heath Stone)
- 4.7 Changement climatique
- 5.0 Publications de 2006-2008
Remerciements
La publication du présent document n'aurait pas été possible sans la contribution et le concours des nombreux chercheurs spécialistes des mammifères marins du ministère des Pêches et des Océans au Canada. Nous remercions le personnel de soutien, les examinateurs et tous ceux qui ont offert une aide précieuse pour la production de cette publication. Nous tenons en outre à remercier Christine Abraham, Estelle Couture et Kerri Swail pour toutes les heures de travail consacrées à l'élaboration du rapport.
Nous aimerions remercier plus particulièrement les personnes suivantes qui ont contribué à la rédaction du texte:
- Don Bowen - Chercheur scientifique, Halifax (N.-É.)
- Larry Dueck - Biologiste, Winnipeg (Man.)
- Steve Ferguson - Chercheur scientifique, Winnipeg (Man.)
- John Ford - Chercheur scientifique, Nanaimo (C.-B.)
- Jean-François Gosselin - Chercheur scientifique, Mont-Joli (QC)
- Mike Hammill - Chercheur scientifique, Mont-Joli (QC)
- Lois Harwood - Biologiste, Yellowknife (T. N.-O.)
- Jeff Higdon - Biologiste, Winnipeg (Man.)
- Jack Lawson - Chercheur scientifique, St. John's (T.-N.)
- Véronique Lesage - Chercheure scientifique, Mont-Joli (QC)
- Lena Measures - Chercheure scientifique, Mont-Joli (QC)
- Linda Nichol - Biologiste, Nanaimo (C.-B.)
- Peter Olesiuk - Biologiste, Nanaimo (C.-B.)
- Lianne Postma - Biologiste, Winnipeg (Man.)
- Pierre Richard - Chercheur scientifique, Winnipeg (Man.)
- Peter Ross - Chercheur scientifique, Sidney (C.-B.)
- Yvan Simard - Chercheur scientifique, Mont-Joli (QC)
- Becky Sjare - Chercheure scientifique, St. John's (T.-N.)
- Kent Smedbol - Chercheur scientifique, St. Andrews (N.-B.)
- Garry Stenson - Chercheur scientifique, St. John's (T.-N.)
- Heath Stone - Biologiste, St. Andrews (N.-B.)
- Edward Trippel - Chercheur scientifique, St. Andrews (N.-B.)
Message du directeur du Centre d'expertise sur les mammifères marins

Mike Hammill, directeur du CEMAM
Photo: Garry Stenson
1.0 Aperçu
Le CEMAM est chargé d'établir les priorités en matière de recherche sur les mammifères marins et de faire mieux connaître son programme sur les mammifères marins. Les scientifiques spécialistes des mammifères marins et le CENAM ont de nombreuses raisons d'effectuer des activités de recherche. Celles-ci font appel à des collaborations nationales et internationales avec l'industrie, le milieu universitaire, des organisations non gouvernementales (ONG) et d'autres organismes gouvernementaux. Par exemple, nombre des mammifères marins de l'Arctique sont gérés conjointement avec le Conseil de gestion des ressources fauniques du Nunavut.
De nombreuses obligations nous incombent en vertu de la Loi sur les espèces en péril, car de nombreux mammifères marins au Canada font partie des espèces menacées ou en danger de disparition. Des avis doivent donc être formulés relativement aux possibilités de rétablissement. Des recherches doivent également être menées à bien conformément à la Loi sur les océans, en vertu de laquelle des zones de protection marine sont créées, et à la Loi sur les pêches, qui renferme un article visant à assurer la protection des mammifères marins. Plusieurs espèces de mammifères marins sont chassées au Canada, à la fois à des fins commerciales et de subsistance. Le Règlement sur les mammifères marins de la Loi sur les pêches a pour but de réglementer et de surveiller ces chasses. Il incombe au CENAM d'effectuer des relevés et des évaluations afin de fournir des estimations de l'abondance et les totaux autorisés de captures pour ces chasses.
Au sein du CEMAM, il existe plusieurs priorités de recherche, notamment la productivité des populations et des communautés de poissons, les liens entre l'habitat et la population, les changements et la variabilité climatiques et la santé des animaux aquatiques. Celles-ci sont liées aux résultats stratégiques qui incluent la compréhension et la description de l'état des écosystèmes aquatiques et l'évaluation et l'atténuation des effets des activités humaines.
Il existe trois grands thèmes de recherche, qui constituent les piliers du présent rapport. Ces trois thèmes de recherche se rapportent aux quatre objectifs du CENAM: la recherche sur la population, le rôle des mammifères marins dans l'écosystème, les impacts du développement et la santé des animaux aquatiques.
La dynamique des populations est l'étude de la taille des populations et des facteurs qui ont un effet sur l'abondance des animaux. Ces facteurs comprennent les taux de reproduction et de mortalité selon l'âge ainsi que les niveaux d'immigration ou d'émigration. Les chercheurs examinent également des facteurs importants tels que l'apport énergétique ou la maladie lorsqu'ils déterminent la dynamique des populations.
Le rôle des mammifères marins dans les écosystèmes marins est le deuxième thème et une question complexe. Il faut disposer de nombreuses données, entre autres sur la consommation globale. Pour évaluer les impacts, il faut obtenir des données sur l'abondance des proies, ainsi que sur l'emplacement des proies, les besoins nutritionnels et la dépense d'énergie. Certains mammifères marins sont des prédateurs d'un niveau trophique supérieur tandis que d'autres sont plus bas dans la chaîne trophique, par exemple ceux qui s'alimentent d'organismes benthiques et d'invertébrés pélagiques.
L'impact de l'homme sur les mammifères marins constitue le troisième et dernier thème. L'impact le plus évident est la capture à des fins commerciales et de subsistance. Les autres impacts sont, sans s'y limiter, l'observation de mammifères marins, l'exploration pétrolière, la navigation commerciale et l'aménagement du littoral. Ceux-ci peuvent avoir des répercussions à court terme ou à long terme sur la survie.
La présente publication porte sur les recherches effectuées ces trois dernières années par les scientifiques du CEMAM, réparties selon les trois piliers. Le but de la publication est de donner un aperçu de la diversité de la recherche réalisée sur les mammifères marins au MPO.
2.0 Dynamique des populations
2.1 Abondance
2.1.1 SPLASH: Étude internationale sur les rorquals à bosse dans le Pacifique Nord
John Ford

Figure 1. Lieux d'observation de plus de 6 000 rorquals à bosse au large des côtes de Colombie-Britannique, 1982-2007.
À l'instar de nombreuses populations mondiales de grandes baleines, la population de rorquals à bosse dans le Pacifique Nord a été décimée par la chasse à la baleine pratiquée du début jusqu'à la moitié du XXe siècle. Quelque 28 000 rorquals à bosse ont été tués dans l'est du Pacifique Nord seulement, et à la fin de la chasse commerciale à la baleine en 1965, il restait environ 2 000 baleines, peut-être moins. Au cours des dernières années, le rorqual à bosse a présenté des signes de rétablissement, mais on ne dispose d'aucune estimation fiable de l'abondance de la population ou des tendances pour le Pacifique Nord. L'espèce est toujours considérée comme une espèce menacée en vertu de la Loi sur les espèces en péril du Canada.
Au début des années 2000, un groupe de chercheurs spécialistes des baleines de plusieurs pays côtiers du Pacifique, y compris le Canada, a proposé une étude ambitieuse pour déterminer la situation du rorqual à bosse dans le Pacifique Nord. Appelée SPLASH (acronyme formé des mots anglais « Structure of Populations, Levels of Abundance and Status of Humpbacks », structure des populations, niveaux d'abondace et situation des rorquals à bosse), cette initiative est finalement devenue la plus grande étude sur les baleines jamais entreprise. Elle a fait appel pendant trois ans à plus de 400 chercheurs de 10 pays (Canada, États-Unis, Russie, Japon, Mexique, Philippines, Guatemala, Nicaragua, El Salvador et Costa Rica). Cette étude sur le terrain reposait principalement sur deux approches: la photo-identification des baleines à l'aide de marqueurs naturels sur la pointe de la queue, pour déterminer l'abondance de la population et les liens migratoires entre les aires de reproduction et les aires d'alimentation, et une analyse de l'ADN de tissus cutanés prélevés pour déterminer la structure génétique de la population.
L'étude SPLASH a donné d'excellents résultats. Plus de 8 000 rorquals à bosse ont été catalogués. Par suite des analyses des marquages et des recaptures de cet ensemble de données, la population a été estimée à environ 18 000 baleines dans le Pacifique Nord. Des échantillons de peau ont été prélevés sur plus de 6 000 individus et ceux-ci ont révélé une structure génétique complexe dans les diverses régions du Pacifique Nord.

Nageoires caudales de rorquals à bosse et navire de la Garde côtière canadienne
Photo: John Ford
Les scientifiques du MPO ont joué un rôle de premier plan dans l'étude SPLASH, depuis les premières étapes de la planification jusqu'à l'analyse de milliers de photos d'identification, en passant par la recherche intensive sur le terrain au large des côtes de Colombie-Britannique (figure 1). Comme dans d'autres zones du Pacifique Nord, la population de rorquals à bosse qui se nourrit au large de la côte Pacifique du Canada croît rapidement à un taux annuel estimatif de 4 % en raison du recrutement et d'un taux élevé de survie proche de près de 98 % par an. Selon une estimation réaliste, l'abondance de rorquals à bosse dans les eaux de la Colombie-Britannique serait d'environ 2 000 individus.
Les photographies de rorquals à bosse prises en Colombie-Britannique au cours de l'été et de l'hiver ont été comparées avec celles prises dans des aires de reproduction subtropicales, où ces baleines se rassemblent en hiver. La comparaison a révélé que les rorquals à bosse de la Colombie-Britannique migrent principalement vers deux destinations différentes: vers les eaux au large de la côte ouest du Mexique dans le cas des individus au large de l'île de Vancouver, tandis que ceux présents au large de la côte nord de la Colombie-Britannique migrent principalement vers Hawaï.
SPLASH a permis de mieux comprendre la situation des rorquals à bosse dans le Pacifique Nord ainsi que dans les eaux canadiennes. Elle est venue fort à propos puisque le MPO élabore actuellement une stratégie de rétablissement du rorqual à bosse dans les eaux du Pacifique canadien. Le rorqual à bosse semble bien se rétablir, même s'il n'est pas encore hors de danger. Les risques pesant actuellement sur la survie et le rétablissement de la population de rorquals à bosse sont entre autres les collisions avec des navires, l'enchevêtrement dans les engins de pêche, l'augmentation du bruit sous-marin et la disponibilité limitée des proies. Toutefois, aucune de ces menaces ne semble nuire actuellement au taux de croissance soutenu de la population.
2.1.2 Abondance du béluga de l'ouest de la baie d'Hudson
Pierre Richard

Attroupement de bélugas
Photo: Véronique Lesage
La population de bélugas de l'ouest de la baie d'Hudson passe l'été dans les eaux côtières du Manitoba; les plus grands regroupements se retrouvent dans les estuaires des rivières Seal et Churchill et du fleuve Nelson. On a établi deux estimations du nombre d'animaux à la surface (23 000 et 25 100 individus) à partir des relevés photographiques aériens et des observations visuelles effectués en 1987.
Des relevés aériens ont été effectués dans l'ouest et le sud de la baie d'Hudson en 2004. On a utilisé à la fois des relevés visuels aériens par échantillonnage en ligne et des relevés photographiques pour estimer le nombre de bélugas, avec une correction pour tenir compte du biais de disponibilité (animaux manqués par les observateurs car ils étaient sous l'eau). La nouvelle estimation s'élevait à 57 300 (IC de 95 % – 37 700-87 100). Les estimations antérieures par suite du relevé, corrigées pour tenir compte du biais de disponibilité, ont donné des chiffres similaires, ce qui donne à penser que le nombre d'individus n'a pas connu de changements perceptibles depuis 1987.
2.1.3 Abondance du béluga de l'estuaire du Saint-Laurent
Jean-François Gosselin

Photographie du revelé de bélugas
Photo: Jack Orr
Les relevés visant à évaluer l'abondance de bélugas dans le Saint-Laurent ont utilisé diverses méthodes, entre autres des bateaux, des hélicoptères, des aéronefs, des observations visuelles et des relevés photographiques. Le plan de rétablissement du béluga du Saint-Laurent de 1995 recommandait l'adoption d'une méthode standard pour évaluer l'abondance et améliorer la surveillance de la population. Depuis 1988, sept relevés des bélugas dans l'estuaire du Saint-Laurent ont utilisé une méthode de relevé photographique aérien systématique. Le dernier de ces relevés a été réalisé en 2003 et après avoir appliqué un facteur de correction pour tenir compte du nombre d'animaux sous l'eau pendant le survol de l'aéronef, l'indice d'abondance s'élevait à 1 300 (arrondi à la centaine près). Utilisant les estimations photographiques établies depuis 1988, un modèle démographique donne à penser qu'en dépit des 30 années de protection contre la chasse, le béluga de l'estuaire du Saint-Laurent ne montre aucun signe de rétablissement, la population comptant environ 1 100 individus (arrondis à la centaine près).
On a observé une variabilité importante entre les indices d'abondance depuis 1988, qui, croit-on, est attribuable aux problèmes rencontrés lorsqu'on essaie d'effectuer un relevé d'une petite population ayant une distribution agrégative. En 2005, on s'est appuyé sur 19 relevés visuels par échantillonnage en ligne effectué pour évaluer l'effet de l'altitude sur les estimations d'abondance utilisant cette méthode, ainsi que cinq relevés visuels par échantillonnage en ligne effectués en 2003 pour établir en partie la variabilité associée à la distribution agrégative des bélugas dans le Saint-Laurent et pour comparer les estimations des deux méthodes de relevés. L'indice d'abondance du relevé photographique de 2003 était inférieur, représentant 68 % de l'estimation combinée des cinq relevés visuels par échantillonnage en ligne de cette même année, mais n'était pas différent des estimations combinées des relevés visuels de 2005. Même si les indices d'abondance des deux méthodes n'étaient pas différents, il faudrait procéder à des comparaisons additionnelles pour assurer la calibration de ces deux techniques.
2.1.4 Structure par âge, croissance, mortalité et densité de la population de bélugas dans l'Arctique canadien
Steve Ferguson

Béluga et son baleineau
Photo: W. Klenner
Les écosystèmes marins polaires et subpolaires affichent les plus grandes variations saisonnières du monde et les plus grands extrêmes dans les caractéristiques physiques de l'environnement. Lorsque l'environnement est à ce point déterminant, les stratégies d'évolution biologique sont souvent considérées comme des adaptations à divers degrés de saisonnalité et/ou de prévisibilité des ressources, en général alimentaires. Les contraintes environnementales devraient croître avec la saisonnalité des ressources, notamment alimentaires, et d'un habitat de reproduction adéquat.
Il est difficile de vérifier les prévisions concernant l'influence des facteurs environnementaux et de la densité en raison de l'envergure limitée des comparaisons de la structure par âge et de la croissance entre les populations de bélugas. Une comparaison détaillée s'impose, puisque deux stocks canadiens sont répertoriés parmi les espèces menacées d'extinction, deux sont considérées comme des espèces menacées et deux font partie des espèces préoccupantes en vertu de la Loi sur les espèces en péril.
Dans cette étude, on a comparé la structure par âge, la répartition selon la longueur corporelle, la mortalité et la densité de quatre populations de bélugas de l'Arctique canadien: baie Baffin, baie Cumberland, ouest de la baie d'Hudson et est de la baie d'Hudson. La croissance, mais non la mortalité, montre une corrélation positive significative avec la latitude. La répartition par âge diffère dans sa forme entre les populations, les animaux évoluant aux latitudes les plus septentrionales (est de la mer de Beaufort) étant les plus vieux et ayant les plus grandes longueurs corporelles à l'âge adulte. Même si les différences dans la sélectivité et la pression de la chasse ont peut-être joué un rôle dans les réponses observées, la variation dans la taille corporelle de l'adulte donne à penser que la saisonnalité environnementale exerce peut-être des contraintes plus fortes sur les caractéristiques du cycle biologique des bélugas, qui augmentent à mesure que l'on progresse vers le nord.
2.1.5 Réduction soutenue de la croissance démographique du phoque gris
Don Bowen et Mike Hammill

Colonie de phoques gris sur la plage nord de l'île de Sable
Photo: Don Bowen
L'une des plus longues séries d'estimations de la production de jeunes phoques gris provient d'une étude amorcée en 1962 à l'île de Sable, en Nouvelle-Écosse. Le nombre de jeunes phoques gris dans l'île de Sable a augmenté de manière exponentielle au cours des quarante dernières années. Dans son aire de distribution au Canada, le phoque gris part à la recherche de nourriture principalement sur la Plateforme Scotian et dans le golfe du Saint-Laurent en dehors de la saison de reproduction. Les plus grandes colonies de reproduction se trouvent sur la banquise dans la partie sud du golfe du Saint-Laurent et sur l'île de Sable.
Un relevé aérien utilisant la photographie numérique a été réalisé dans l'île de Sable en janvier 2007 pour estimer la production de jeunes phoques gris. On a dénombré un total de 48 036 nouveau-nés sur les photographies numériques. Après correction pour tenir compte de la proportion de nouveau-nés morts avant le relevé et de ceux nés après le relevé, la nouvelle estimation était de 54 500 individus. Pour le segment de population ne vivant pas sur l'île de Sable, des relevés aériens visuels par bande ont été réalisés pour les concentrations de mise bas sur la glace du golfe du Saint-Laurent et des dénombrements ont été réalisés sur les îles du golfe et le long de la côte est de la Nouvelle-Écosse. Des recherches aériennes ont également été effectuées le long de la côte sud et sud-ouest de Terre-Neuve et de l'île d'Anticosti. Les dénombrements ont été corrigés pour tenir compte de la proportion de mises bas qui a eu lieu après la réalisation du relevé.
Dans la mesure du possible, les dénombrements ont été ajustés en fonction de la proportion de mises bas qui a eu lieu après la réalisation du relevé. Les changements dans la durée de l'étape du pelage propre à l'âge ont donné lieu à un changement de 17 % dans les estimations de production de nouveau-nés. La production totale de nouveau-nés en dehors de l'île de Sable, arrondie au millier près, est estimée à 13 000 animaux. La proportion de jeunes nés sur l'île de Sable a augmenté, passant de moins de 5 % vers le milieu des années 1980 à environ 70 % en 2007.
Selon l'estimation de 2007, la production de nouveau-nés sur l'île de Sable a continué d'augmenter, mais le taux d'accroissement a baissé au cours des deux derniers relevés. La proportion de femelles mettant bas à l'âge de 4 à 7 ans était bien moins grande pour les cohortes de 1998-2002 que pour celle observée de la moitié à la fin des années 1980. La nouvelle estimation de la production de nouveau-nés et les nouvelles données sur l'âge de la première mise bas donnent de nouvelles indications du changement dans les indices vitaux de cette population. La production de nouveau-nés en dehors de l'île de Sable n'a pas affiché la même augmentation que celle observée sur l'île de Sable. Ceci est peut-être dû à des taux de mortalité plus élevés et plus variables des jeunes nés sur la glace dans le golfe du Saint-Laurent.
Les données du relevé ont été intégrées à un modèle démographique en même temps que des données sur les taux de reproduction et sur les prises déclarées. La taille totale estimative de la population à la fin de la saison de reproduction 2007 (c'est-à-dire incluant les nouveau-nés) s'élevait à 304 000 individus.
2.1.6 Moment de la mise bas et abondance des phoques à capuchon dans l'Atlantique Nord-Ouest
Garry Stenson et Mike Hammill

Phoque à capuchon mâle
Photo: Mike Hammill
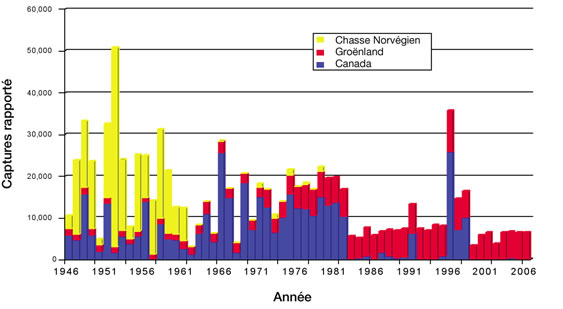
Figure 2. Prises déclarées de phoques à capuchon dans l'Atlantique Nord-Ouest, 1946-2006.
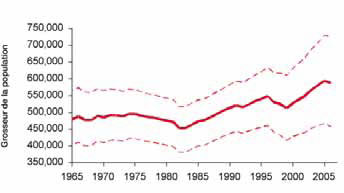
Figure 3. Abondance estimative du phoque à capuchon dans l'Atlantique Nord-Ouest, 1865-2006.
La mise bas des phoques à capuchon a lieu sur la banquise autour de l'île Jan Mayen, dans le détroit de Davis, au large de la côte nord-est de Terre-Neuve (appelée le Front) et dans le golfe du Saint-Laurent. Les relations entre les différents groupes reproducteurs sont mal comprises; certaines analyses utilisant des techniques d'analyse de l'ADN appuient l'hypothèse de l'existence d'une seule population.
Des relevés ont été effectués pendant la saison de reproduction afin d'étudier le moment de la mise bas au printemps. Au Front, la date moyenne de fin de la mise bas était le 28 mars, soit entre le 18 mars et le 4 avril. Dans le golfe, la mise bas prend fin presque tous les ans avant le début d'avril, sauf en 1994, où le modèle a prédit que la mise bas se poursuivrait jusqu'en mai. Si l'on exclut ces données, les naissances auraient pris fin le 28 mars, soit au plus tôt le 25 mars et au plus tard le 31 mars.
On a observé des phoques à capuchon femelles allaitant leurs petits au début mars, ce qui indique que la mise bas de ces phoques s'étale sur une plus longue période que ce que l'on a observé chez les phoques du Groenland qui mettent bas dans la même région. Si l'on tient compte de la possibilité de naissances tardives, en particulier pendant les années de glace dense, l'allaitement devrait être terminé vers le 8-10 avril.
Utilisant des analyses séquentielles des populations ou des indices de survie, les chercheurs ont estimé que la production de jeunes phoques à capuchon au Front était d'environ 30 000 individus au début des années 1980. Toutefois, selon des estimations de la production de jeunes établies grâce à des relevés aériens indépendants effectués au milieu des années 1980, la production de jeunes était d'environ 62 000 pendant les années 1980 et de 83 000 en 1990. Malheureusement, on ne connaît pas bien l'abondance historique des animaux dans le détroit de Davis et dans le golfe. Selon un relevé effectué en 1984, la production estimative de nouveau-nés dans le détroit de Davis serait d'environ 18 600 individus. Relativement peu de jeunes sont nés dans le golfe du Saint-Laurent, les estimations des relevés aériens se chiffrant à 2 000 et à 4 000 animaux en 1990 et 1994 respectivement.
Les estimations les plus récentes de la production de nouveau-nés proviennent des relevés du Front effectués en 2004 et des trois aires de mise bas réalisés en 2005. En 2004, la production de nouveau-nés au Front a été estimée à 123 862. En 2005, elle était estimée à 107 013, tandis qu'on estimait que 6 620 jeunes étaient nés dans le golfe. La production de nouveau-nés dans la concentration de mise bas dans le détroit de Davis était estimée à 3 346. Si l'on combine ces régions, on obtient une production estimative de nouveau-nés pour les trois aires de mise bas de l'Atlantique Nord-Ouest s'élevant à 116 900 individus. La comparaison avec des estimations antérieures donne à penser que la production de jeunes a peut-être augmenté depuis le milieu des années 1980.
Un modèle démographique incorporant les estimations de production des jeunes phoques à capuchon depuis les années 1980, les taux de reproduction et la mortalité causée par l'homme (prises déclarées et individus abattus et perdus) a été utilisé pour estimer l'abondance totale pour la période 1965-2005 (figure 2).
Fitting to pup production estimates from all herds and making assumptions about numbers of hooded seals in the Davis Strait herd for years when this area was not included in the survey program, resulted in an estimated total population of 593,500 in 2006 (Fig. 3). However, there is considerable uncertainty associated with these estimates owing to our lack of understanding of the relationship between the Davis Strait, Front and Gulf pupping areas, few surveys of all three areas, limited reproductive data and uncertain harvest statistics.
2.1.7 Abondance de l'otarie de Steller en Colombie-Britannique
Peter Olesiuk

Abondance de l'otarie de Steller en Colombie-Britannique
Photo: croisière de recherche UBC-MPO
Les otaries de Steller se reproduisent le long du littoral du Pacifique Nord. On connaît deux populations distinctes: une population de l'ouest qui se reproduit en Russie, dans les îles Aléoutiennes et dans le golfe de l'Alaska, et une population de l'est, qui se reproduit dans le sud-est de l'Alaska, en Colombie-Britannique, dans l'Oregon et dans le centre-nord de la Californie. Le MPO effectue des relevés aériens à intervalles de deux à cinq ans depuis le début des années 1970 afin de surveiller les populations de la Colombie-Britannique. Depuis 1994, les relevés du MPO ont été coordonnés avec ceux d'organismes de l'Alaska, de Washington, de l'Oregon, de la Californie et de la Russie afin d'obtenir des estimations sur l'abondance pour l'ensemble de l'aire de répartition de l'espèce.
Les relevés ont lieu entre la fin de juin et le début de juillet, afin de coïncider avec la fin de la saison de mise bas. La plupart des jeunes sont alors nés, mais ils sont encore trop jeunes pour avoir quitté les roqueries, si bien que les relevés fournissent une estimation de la production totale annuelle de nouveau-nés. Des levés des échoueries utilisées par des animaux non reproducteurs sont également effectués. Même si certains individus autres que des jeunes se trouvent en mer et ne sont pas comptabilisés pendant les relevés, ces derniers fournissent de l'information sur la répartition relative des animaux et l'importance des échoueries.
Selon les relevés effectués en Colombie-Britannique, la tendance est à la hausse tant pour ce qui est de la production de jeunes que des dénombrements d'individus autres que des jeunes depuis le début des années 1970, la plupart des augmentations ayant eu lieu depuis les années 1980. L'abondance de l'otarie de Steller a également augmenté dans le sud-est de l'Alaska, l'État de Washington et l'Oregon. Comme les plus grandes roqueries se trouvent près de la frontière entre la Colombie-Britannique et le sud-est de l'Alaska, il est difficile de séparer les populations de ces deux régions, mais toutes deux affichent des tendances à la hausse. En fait, l'abondance combinée des otaries de Steller en Colombie-Britannique et dans le sud-est de l'Alaska a presque quadruplé depuis que l'espèce est protégée, soit depuis le début des années 1970.
Même si des relevés n'ont pas été réalisés à l'échelle de la Colombie-Britannique avant le début des années 1970, il existe des dénombrements pour des roqueries de reproduction datant du début des années 1900. Selon des relevés récents, une proportion relativement constante de la population totale se trouve sur les roqueries, si bien que les dénombrements historiques qui y sont effectués constituent probablement un indice relativement fiable des tendances démographiques globales. Les dénombrements historiques indiquent que l'abondance des otaries en Colombie-Britannique au début des années 1970 a diminué pour s'établir à environ 25-33 % du nombre observé quand les premiers relevés ont été effectués en 1913. Les baisses sont attribuables aux programmes de contrôle des prédateurs et aux pêches expérimentales. Environ 55 000 otaries de Steller ont été tuées en Colombie-Britannique entre 1912 et 1968, ce qui a éliminé une roquerie dans l'archipel Sea Otter Group au large des côtes du centre de la Colombie-Britannique, et réduit les nombres sur les trois roqueries restantes.
Les récentes augmentations en Colombie-Britannique et dans les eaux adjacentes reflètent le rétablissement de populations qui avaient été décimées avant d'être protégées au début des années 1970. L'espèce a recommencé à se reproduire dans l'archipel Sea Otter Group et en 2006, un nombre suffisant de jeunes y sont nés pour qu'on puisse à nouveau la qualifier de roquerie. Ces dernières années, les populations d'otaries de Steller en Colombie-Britannique et dans le sud-est de l'Alaska semblent avoir dépassé les niveaux record historiques et on ne connaît pas la raison de cette augmentation soutenue. En 2002, la population de l'est totale des otaries de Steller était estimée à 46 000-58 000, dont 34 % se reproduisent sur la côte de la Colombie-Britannique.
2.1.8 Abondance des baleines boréales de l'est de l'Arctique
Larry Dueck

Abondance des baleines boréales de l'est de l'Arctique
Photo: Larry Dueck
Il est reconnu que l'abondance des baleines boréales soit difficile à évaluer. Outre leur répartition très étendue et leur densité relativement faible, elles ont aussi tendance à échapper à la vue, sous la surface de l'eau, et sont souvent difficiles à distinguer en raison de leur couleur foncée. Il faut effectuer des relevés dans un délai aussi court que possible, et les conditions atmosphériques et de visibilité compliquent encore la tâche, si bien que des parties importantes du secteur du relevé sont en bout de ligne omis.
Les relevés antérieurs de baleines boréales dans l'est de l'Arctique ne couvraient que des petites parties de leur aire de distribution en raison du coût des relevés aériens et avaient tendance à grandement différer, principalement en raison de facteurs liés aux conditions du relevé et au territoire couvert. Les premiers relevés ont également sous-estimé le nombre de baleines, puisqu'ils ne corrigeaient généralement pas les données pour tenir compte des animaux sous l'eau. L'information sur le comportement de la baleine boréale, en particulier le temps passé sous l'eau, n'était pas encore connue à l'époque.
Une importante opération de relevé a pu être réalisée en 2002-2004 grâce au financement du programme des espèces en péril. Pour obtenir la meilleure information qui soit, on a effectué des relevés aériens à partir d'un aéronef Twin Otter, avec deux observateurs de chaque côté de l'appareil. Cette méthode permet une estimation plus complète du nombre d'animaux puisqu'elle permet aux chercheurs de documenter les espèces de baleines aperçues par l'observateur installé à l'avant, mais non par l'observateur installé à l'arrière, et vice versa. La détermination de la proportion d'animaux non repérés (parce qu'ils se trouvaient sous l'eau) est essentielle pour calculer l'abondance des baleines.
L'analyse de ces données, y compris les ajustements pour tenir compte des animaux passés inaperçus et des animaux sous l'eau, indique que l'abondance des baleines boréales dans l'est de l'Arctique se situe entre 4 800 et 43 000 individus, soit un chiffre estimatif moyen de 14 400 baleines. Même si on ne sait pas précisément où se situe le nombre exact à l'intérieur de cette fourchette, l'estimation nous indique que les nombres révèlent des quantités plus élevées qu'on ne le croyait auparavant et vient corroborer les affirmations des Inuits de nombreuses collectivités selon lesquelles leur nombre a augmenté au cours des dernières décennies.
Note en bas de page: Selon un récent examen du Sous-comité scientifique de la Commission baleinière internationale, l'estimation ci-dessus est peut-être par trop optimiste. Un examen est en cours en vue de répondre à ces préoccupations.
2.2 Établissement de la structure de la population à l'aide de la génétique moléculaire
2.2.1 Différencier les bélugas de l'est de la baie d'Hudson de ceux de l'ouest de la baie d'Hudson au moyen de marqueurs moléculaires
Lianne Postma
L'identification des stocks de bélugas dans la baie d'Hudson et ailleurs dans les eaux canadiennes repose fortement sur l'information génétique. Cette information a été utilisée pour établir une distinction entre les rassemblements estivaux de bélugas appartenant à des lignées maternelles communes. Les analyses préliminaires de ces données corroboraient l'existence d'au moins deux stocks de bélugas dans la baie d'Hudson, généralement appelés stocks de l'est et de l'ouest de la baie d'Hudson. On pense que ces stocks se croisent entre eux dans les aires d'hivernage dans l'est du détroit d'Hudson et dans la baie de Baffin.

Examen des profils d'ADN en laboratoire
Photo: Denise Tenkula, Melissa Lindsay et Robert Bajno
Des échantillons prélevés sur les prises locales ont révélé un mélange d'animaux en migration et d'animaux en estivage. La première étape consiste à relever le nombre de stocks dans la population en isolant les échantillons estivaux. L'étape suivante consiste à examiner les prélèvements des animaux en migration et à effectuer une analyse du stock mélangé pour quantifier les proportions d'animaux des stocks estivaux qui composaient les prises.
Selon les résultats préliminaires de ces analyses, il existe plus de deux stocks de bélugas dans la baie d'Hudson. Tout porte aussi à croire que les échantillons prélevés dans la baie James représentent une population distincte de bélugas qui ne se croisent pas avec les bélugas capturés à d'autres endroits dans la baie d'Hudson et le détroit d'Hudson.
2.2.2 La génétique moléculaire pour l'identification des stocks de narvals au Nunavut
Lianne Postma

Petit groupe de narvals en eau libre
Photo: Larry Dueck
Les résultats des analyses génétiques d'échantillons prélevés sur des narvals dans l'est de l'Arctique canadien entre 1982 et 2001 ont révélé que la différenciation des stocks de narvals dans l'Extrême-Arctique était faible. Il ressort également que le choix des marqueurs aurait dû être suffisant pour mettre au jour les différences entre les stocks.
Dans le cadre des récents travaux concernant le béluga et le morse, on s'est attaché principalement à accroître la quantité d'information génétique utilisée pour examiner les différences entre les stocks et les comportements migratoires. Ces efforts ont accru la robustesse des analyses statistiques et nous ont beaucoup éclairés sur ces espèces.
Au cours des dernières années, les approches analytiques ont grandement changé. Des méthodes plus puissantes récemment mises au point permettent d'établir de manière plus rigoureuse la structure de la population, de quantifier le flux génétique, de fournir de l'information sur l'importance de la migration actuelle par rapport aux associations historiques entre les populations ainsi que de l'information démographique sur l'historique, la croissance et la variabilité de la population. Ces types d'information sont essentiels pour l'établissement de plans de gestion et de plans de rétablissement des stocks en danger de disparition.
Ces méthodes ont toutefois des limites, dont la grande puissance de traitement et la nécessité de disposer de grands ensembles de données. Dans ce projet, le nombre d'échantillons pertinents analysés a été porté à 20 marqueurs microsatellites pour accroître la quantité d'information génétique disponible. L'accent est mis sur l'analyse de 50 échantillons provenant de chacun des quatre lieux – Repulse Bay, Inlet Pond, Fjord Grise et Kikitarjuaq. Les résultats de cette étude sont attendus.
2.2.3 Modes de différenciation génétique chez les baleines boréales de l'est de l'Arctique canadien et de l'ouest du Groenland
Lianne Postma

Chercheurs du MPO marquant des baleines boréales à partir d'un bateau
Photo: DFO
Des échantillons de biopsie ont été prélevés sur des baleines boréales au Nunavut, au Canada, et dans l'ouest du Groenland, et ont été analysés pour 34 loci de microsatellites d'ADN nucléaire. La taille des échantillons était de 8 pour Pelly Bay, 143 pour Igloolik, 16 pour Repulse Bay, 82 pour Pangnirtung et 80 pour Disko Bay.
Les résultats peuvent indiquer la structure des stocks, mais ils pourraient aussi être attribuables à plusieurs facteurs, notamment la consanguinité, le croisement non aléatoire, la sélection naturelle, les conséquences dues à la forte baisse de la population en raison de la chasse commerciale, ou les erreurs de notation des données. Une analyse de la variance moléculaire (AMOVA) a révélé des différences significatives de fréquences d'allèles entre les échantillons d'années différentes à chaque endroit. Ces résultats sont très probablement influencés par la taille des échantillons et la variation dans les sexes et les âges des échantillons de la collection d'une année à l'autre.
La preuve génétique moléculaire des analyses des microsatellites des échantillons de baleines boréales provenant de la baie de Baffin-détroit de Davis et de la baie d'Hudson-bassin Foxe ne semble pas confirmer l'existence de deux populations distinctes. Cette conclusion est corroborée par les résultats des études de localisation par satellite montrant les importants déplacements de baleines boréales entre ces deux régions. Toutefois, les différences génétiques qui ont été relevées et l'information limitée sur la ségrégation en fonction de l'âge et du sexe entre les lieux d'échantillonnage révèlent l'existence d'une certaine structuration des baleines boréales dans leur aire de distribution dont il faudrait peut-être tenir compte dans la gestion de cette population.
2.3 Reproduction et mortalité
2.3.1 Tactique d'accouplement des phoques gris mâles selon leur condition: l'importance de la taille du corps
Don Bowen

Combat de phoques gris mâles sans gagnant
Photo: Don Bowen
Les phoques gris ont une taille dimorphe, les mâles étant 1,5 fois plus grands que les femelles. Chez les espèces dimorphes, on pense souvent que les grands mâles possèdent un avantage pour obtenir une femelle. Cette étude examinait l'importance de la taille du corps et de la composition du corps en tant qu'éléments déterminants de la tactique d'accouplement adoptée par les mâles. Les observations du comportement, ainsi que les mesures de la dépense d'énergie et du succès de reproduction, ont été effectuées pour 42 individus dont on connaissait l'âge pendant les saisons de reproduction de 1997-2001 sur l'île de Sable, au Canada.
Les mâles ayant une masse corporelle élevée arrivaient sur les lieux de reproduction avec plus de réserves lipidiques et d'énergie et dépensaient moins de protéines corporelles que leurs semblables plus petits. Les mâles ayant un pourcentage moins élevé de réserves lipidiques avaient des taux plus élevés d'absorption d'aliments. On observe une corrélation positive entre la quantité d'énergie corporelle à l'arrivée et la durée de la période d'accouplement. Les mâles qui adoptaient pour principale tactique d'accouplement la défense des femelles étaient plus lourds, ils avaient indubitablement plus de tissus adipeux et d'énergie corporelle et ils s'adonnaient plus longtemps à l'accouplement que les mâles qui n'avaient pas adopté cette tactique. Parmi tous les mâles, ceux de taille intermédiaire s'accouplaient avec un plus grand nombre de femelles et obtenaient le plus haut taux de succès. La taille intermédiaire du corps pourrait bien être optimale pendant l'accouplement en raison d'une plus grande agilité durant le combat entre mâles.
2.3.2 Évaluation du dénombrement de nouveau-nés d'otaries de Steller à partir de photographies 35 mm prises à angle oblique
Peter Olesiuk

Les roqueries d'otaries de Steller sont photographiées pendant que l'avion tourne en rond à un angle abrupt et les petits nouveau-nés foncés sont dénombrés à partir de photos haute résolution 35 mm
Photo: Peter Olesiuk
Pour de nombreux pinnipèdes – et l'otarie de Steller ne fait pas exception –, le dénombrement des nouveau-nés constitue la meilleure méthode pour suivre la tendance démographique et estimer l'abondance. La répartition d'autres classes d'âge a tendance à être fort étendue, et une proportion inconnue est en mer et n'est pas comptabilisée pendant les relevés. Par contre, les petits naissent sur les roqueries traditionnelles et sont confinés à terre pendant le premier mois de leur vie. Les recensements de sites de reproduction peuvent donc fournir des estimations de la production totale de nouveau-nés. À partir de là, des modèles démographiques sont utilisés pour extrapoler la taille totale de la population.
Les nouveau-nés d'otaries de Steller sont difficiles à dénombrer à partir de bateaux ou d'avions. Foncés et petits, ils ont tendance à se fondre avec le substrat, et peuvent être dissimulés derrière des rochers, d'autres animaux ou dans des crevasses. Par le passé, on effectuait les dénombrements de nouveau-nés en faisant débarquer des gens sur les roqueries en pleine saison de reproduction, en éloignant les juvéniles et les adultes de la roquerie et en parcourant la roquerie pour compter le nombre de petits présents. Ces dénombrements au sol causent de très fortes perturbations, déplaçant les mâles de leur territoire de reproduction et séparant les petits de leur mère ou les repoussant vers la mer. D'après une étude qui surveillait les changements dans la roquerie avant et après les dénombrements, un tiers de femelles n'ont pas maintenu le contact avec les petits après la perturbation.
En Colombie-Britannique, on a eu recours à des relevés aériens pour surveiller les tendances touchant les otaries de Steller depuis les années 1970. On a dénombré les nouveau-nés à partir de photographies 35 mm prises à angle oblique à partir d'un aéronef qui tournait en rond au-dessus de la roquerie. Si les relevés aériens réduisent la perturbation associée aux dénombrements au sol, on s'interroge néanmoins sur la précision des dénombrements ainsi effectués. Dans les années 1990, les chercheurs du MPO ont fait équipe avec des scientifiques de l'Alaska Department of Fish and Game (ADFG), du National Marine Mammal Laboratory et du Southwest Fisheries Science Center afin de comparer et de calibrer les techniques de dénombrement des petits d'otaries de Steller. Entre 1994 et 1998, le MPO a effectué des relevés à l'île Forrester dans le sud-est de l'Alaska – où naissent plus de nouveau-nés que partout ailleurs – un jour ou deux avant que l'ADFG n'effectue le dénombrement au sol. Si l'on en croit la comparaison, malgré l'existence d'une corrélation étroite entre les relevés aériens et les relevés au sol, invariablement, environ 20 % des petits n'avaient pas été dénombrés par les photographies 35 mm prises à angle oblique.
Dans une étude subséquente, on a constaté que les photographies verticales de format moyen prises à partir d'appareils-photos installés sous l'appareil fournissent des dénombrements statistiquement indifférenciables des dénombrements au sol. En 1998 et en 2002, un relevé des roqueries de la Colombie-Britannique a donc été effectué à l'aide de photographies verticales de format moyen et de photographies 35 mm prises à angle oblique. On observe une corrélation étroite entre les dénombrements utilisant chacune de ces méthodes, mais un petit nombre (4 %) de petits n'ont pas été comptabilisés dans les photographies prises à angle oblique. On croit qu'une proportion plus petite de nouveau-nés étaient dissimulés et n'apparaissaient pas sur les photographies prises à angle oblique des roqueries de la Colombie-Britannique par rapport celles de l'île Forrester du fait que les sites de reproduction de la Colombie-Britannique semblent plus petits ou s'étendre le long des plages, si bien qu'on peut les survoler et les photographier à des angles plus abrupts. Selon ces études, on applique maintenant un petit facteur de correction (1,05) aux dénombrements de jeunes par photographies 35 mm prises à angle oblique en Colombie-Britannique pour tenir compte des animaux dissimulés. Tous les organismes d'Amérique du Nord ont maintenant adopté la photographie aérienne pour le recensement des nouveau-nés d'otaries de Steller, ce qui réduit grandement les perturbations causées par les chercheurs.
2.3.3 Cycle biologique et dynamique des populations d'épaulards résidents du nord de la Colombie-Britannique
Peter Olesiuk, Graeme Ellis and John Ford

Les épaulards résidents demeurent dans leurs groupes natals à vie et se déplacent tous ensemble
Photo: John Ford
Les études sur les populations d'épaulards et les techniques de photo-identification ont été introduites au début des années 1970 par le regretté Michael A. Bigg. Sceptique au départ, Mike a rapidement compris que chaque baleine pourrait être reconnue grâce aux marques naturelles qui lui étaient propres et pris conscience que la photo-identification constituait un puissant outil pour les études démographiques à long terme. Avant son décès prématuré en 1990, il nous a laissé deux documents d'importance capitale qui allaient tracer la voie pour la prochaine génération de chercheurs spécialistes des baleines. Le premier document sur l'organisation sociale et la généalogie montraient que les épaulards résidents des deux sexes demeurent dans leur groupe natal leur vie durant et décrivaient les relations généalogiques entre les individus de chaque groupe matrilinéaire d'épaulards résidents sur la côte de la Colombie-Britannique.
Ce document a ouvert la voie au deuxième document sur le cycle biologique et la biologie des populations d'épaulards résidents. À l'aide de techniques numériques perfectionnées, fort semblables à celles qu'utiliserait un actuaire pour calculer les risques d'assurance, on a établi les paramètres du cycle biologique propres à l'âge d'après les dossiers d'observation de longue date et on les a intégrés aux tables de survie et aux modèles de projection matricielle. Les modèles indiquaient que les épaulards vivaient plus longtemps et se reproduisaient plus lentement que ce que l'on croyait auparavant. Les femelles atteignent généralement la maturité à 14 ans, produisent en moyenne 5 baleineaux sur une durée de vie reproductive de 25 ans, mais peuvent vivre jusqu'à 80 ans. Les mâles atteignent la maturité sexuelle à 13 ans, mais ne sont pas physiquement adultes avant 18 ans, et peuvent vivre jusqu'à 60-70 ans.
On a continué à effectuer des relevés par photo-identification chaque année, et les données sur les naissances et la mortalité des populations d'épaulards résidents couvrent maintenant plus de trente ans. Cette étude de longue haleine nous a offert la possibilité exceptionnelle d'évaluer la façon dont les épaulards s'en sont tirés tant pendant les périodes favorables de croissance démographique que pendant les périodes moins favorables de déclin de la population. Il est intéressant de noter que la situation démographique dépend principalement des changements dans les taux de survie, les taux de reproduction demeurant relativement stables. La stabilité des paramètres de reproduction est peut-être due au potentiel de reproduction très limité des femelles, qui ne produisent qu'en moyenne 5 baleineaux au cours de leur vie. Plus intéressant encore, les animaux des deux sexes et de tous les âges ont affiché des taux de mortalité élevés pendant des périodes moins favorables où les populations déclinaient. Chez la plupart des autres espèces, les jeunes et les animaux inexpérimentés, ou les animaux âgés et frêles sont les plus vulnérables pendant des périodes moins favorables. Cette tendance chez les épaulards peut s'expliquer du fait que, contrairement à d'autres espèces, les animaux des deux sexes et de tous les âges se déplacent ensemble dans leur groupe natal, connaissent les mêmes conditions et souffrent du même manque de proies.
2.4 Rôle des maladies dans les populations de mammifères marins
2.4.1 Maladies importantes pour les populations sauvages de mammifères marins
Lena Measures

Diapositive sur l'histologie de la maladie de Carré du furet
Photo: Lena Measures
Les agents pathogènes les plus importants à l'origine de mortalités massives dans les populations de mammifères marins à travers le monde sont des morbillivirus comme le virus de la maladie de Carré (VMC) infectant les pinnipèdes et le morbillivirus des cétacés (MVC) infectant les cétacés. Le VMC a été responsable de la mort d'environ 20 000 phoques communs et de plusieurs centaines de phoques gris en Europe en 1987-1988 et en 2002. Le MVC a tué des dauphins en Europe et dans l'est des États-Unis, mais le nombre de morts semble plus limité. Le VMC est répandu chez les pinnipèdes canadiens, notamment les phoques du Groenland et les phoques gris, qui seraient les réservoirs du virus. L'absence de mortalité à grande échelle parmi les populations canadiennes de pinnipèdes serait due à l'immunité naturelle du troupeau.
On a signalé quelques cas de mortalité dus au VMC parmi les phoques communs dans la région de la Nouvelle-Angleterre, les plus récents remontant à 2006-2007. On a rapporté la mort d'un seul phoque du Groenland attribuable au VMC dans les eaux canadiennes. Des anticorps contre le VMC avaient été décelés chez les pinnipèdes canadiens dès 1972, mais le virus n'a pas été isolé chez les pinnipèdes canadiens malgré de récents efforts, et aucune étude sur sa pathogénicité ou sa pathogénie n'a été réalisée chez les phoques du Groenland ou les phoques gris. Des études effectuées récemment par Ole Nielsen et Lena Measures du MPO, et des collaborateurs du Centre national des maladies animales exotiques, de l'Agence canadienne d'inspection des aliments et de l'Université de Montréal ont examiné une nouvelle méthode pour isoler et caractériser un isolat de VMC d'une épizootie du phoque survenue en 1988 en Europe en utilisant des furets infectés à des fins expérimentales. Une lignée cellulaire Vero transfectée de manière stable (Vero.DogSLAMtag) exprimant les « molécules canines induisant des signaux d'activation dans les lymphocytes » (SLAM) ou des récepteurs CD150 offrait une réplication virale plus rapide, ce qui a donné lieu à un isolement primaire du virus dans un délai plus court, avec une plus grande production de virus par rapport aux méthodes traditionnelles de culture cellulaire (cellules rénales primaires du phoque et cellules Vero).
Une méthode d'amplification en chaîne par polymérase avec transcription inverse a été mise au point pour déceler une partie du gène du capside nucléique présent uniquement dans le VMC. Cette méthode a été utilisée pour confirmer l'identité du virus qui a par la suite été isolé chez les furets. Les isolats viraux des furets infectés ainsi que les virus isolés au départ chez un dauphin et un marsouin et maintenus dans des cellules Vero se sont aussi reproduits plus vite et ont produit des titres plus élevés de virus lorsqu'ils se sont propagés dans les cellules Vero.DogSLAMtag.
2.4.2 Zoonotic Diseases of Marine Mammals
Lena Measures

Trophozoïtes Giardia
Photo: L. Measures and A. Applebee
Giardia et Cryptosporidium, parasites protozoaires que l'on sait à l'origine des maladies entériques chez les espèces sauvages terrestres, sont également zoonotiques, c'est-à-dire capables d'infecter les humains. De récentes études ont révélé la présence de kystes de Giardia dans les fèces de pinnipèdes canadiens de l'ouest de l'Arctique et de la côte est du Canada. L'objectif de cette nouvelle étude menée à l'Institut Maurice-Lamontagne par Lena Measures et Amber Applebee, dans le cadre de ses recherches doctorales à l'Université Murdoch, à Perth, en Australie, visait à déterminer la prévalence et le génotype de Giardia et Cryptosporidium dans deux espèces de pinnipèdes, le phoque du Groenland (Phoca groenlandica) et le phoque à capuchon (Cystophora cristata), du golfe du Saint-Laurent.
Des échantillons de fèces ont été prélevés sur des phoques adultes et nouveau-nés et examinés en vue de déceler la présence de kystes de Giardia et d'oocystes de Cryptosporidium à l'aide de la microscopie et de la coloration par immunofluorescence. Des tissus du petit intestin des phoques adultes ont été prélevés et examinés en vue de déceler des infections à l'aide de la technique d'amplification en chaîne par polymérase (ACP). On a trouvé des kystes de Giardia dans les fèces de 42 % (16/38) des phoques du Groenland adultes, mais dans aucuns des nouveau-nés de cette espèce (0/20). Même si l'on n'a pas décelé de kystes de Giardia dans les fèces des phoques à capuchon adultes (0/10) à l'aide de la microscopie, 80 % de leurs tissus intestinaux étaient infectés par Giardia d'après la technique d'ACP.
Tant les phoques du Groenland que les phoques à capuchon ont présentés des infections par la souche zoonotique, Giardia duodenalis (assemblage A), déterminée à l'aide d'une technique d'ACP par amorces incluses en deux étapes pour amplifier le gène codant pour la petite sous-unité d'ARNr Giardia. La présence de Cryptosporidium n'a été décelée à l'aide de la microscopie ou de la technique d'ACP dans aucun des tissus intestinaux prélevés des 68 phoques examinés. On a observé des trophozoïtes de Giardia dans les sections histologiques de tissus intestinaux de phoques du Groenland et à capuchon confirmant le dékystement et la prolifération de Giardia chez les phoques.
2.4.3 Échouages et événements de mortalité chez les mammifères marins – Surveillance et enquêtes
Lena Measures

Globicéphale noir vivant échoué
Photo: Lena Measures

Nécropsie d'un béluga
Photo: Lena Measures
Un programme de surveillance et de nécropsie visant la population menacée de bélugas de l'estuaire du Saint-Laurent (ESL) est mené depuis 1982. On signale en moyenne 15 mortalités chaque année, entre trois et douze carcasses étant transportées à la faculté de médecine vétérinaire de l'Université de Montréal pour qu'y soit pratiquée une nécropsie complète en vue de déterminer la cause de la mort et de prélever des tissus pour divers projets de recherche portant sur la santé, le régime alimentaire et les contaminants chimiques présents dans les animaux échoués de cette population. En 2006 et 2007, on a documenté 11 et 17 mortalités, avec 3 et 10 nécropsies réalisées, respectivement.
Des travaux récents effectués avec Michel Lebeuf au MPO ont montré que certains contaminants présents dans les tissus des bélugas de l'ESL diminuaient, tandis que de nouveaux étaient décelés. Entre 1983 et 2007, on a documenté 365 mortalités. Sur les 148 carcasses (les deux sexes et tous les âges confondus) suffisamment fraîches pour la nécropsie entre 1983 et 2002, on a constaté que l'étiologie infectieuse (infections bactériennes et parasitaires) a causé 38 % des décès, la néoplasie ou le cancer terminal 15 %, et que près de 30 % sont morts de causes inconnues. En stratifiant les groupes d'âge, 58 % des baleineaux (moins d'un an) sont morts à la naissance ou juste après la naissance, 81 % des juvéniles (1 à 10 ou 14 ans, selon le sexe) sont morts d'une maladie infectieuse (56 % en raison d'une pneumonie vermineuse), tout comme 32 % des adultes (plus de 10 ou 14 ans selon le sexe). La néoplasie terminale n'a été observée que chez 22 adultes (18 %), dont la moyenne d'âge était de 33 ans (fourchette de 22-59) (GCC/1 an).
On trouvera un aperçu de la population de bélugas de l'ESL intitulé « Quelles sont les causes de mortalité des bélugas du Saint-Laurent? » dans les articles vedettes des Sciences du MPO à l'adresse http://www.dfo-mpo.gc.ca/science/Publications/article/2007/27-07-2007-fra.htm. Malgré la mise en œuvre de diverses mesures de gestion protectrice, l'absence apparente de prédateurs comme les épaulards et l'application de règlements environnementaux sur les produits chimiques toxiques, la population ne s'est pas rétablie. Cet article porte sur l'importance des maladies dans cette population, en particulier les maladies chroniques comme le cancer touchant des animaux qui peuvent vivre plus de 80 ans.
3.0 La place des mammifères marins dans l'écosystème
3.1 Répartition
3.1.1 Information sur l'épaulard de l'Arctique provenant du savoir écologique ancestral et des réseaux d'observation
Steve Ferguson

Épaulard mâle
Photo: John Ford
Le nombre d'épaulards observés dans l'est de l'Arctique canadien a augmenté au cours des dernières années, en particulier dans l'ouest de la baie d'Hudson, et leur répartition a progressé avec les changements climatiques. Toutefois, on en sait peu sur leur répartition et leur écologie.
Le projet Orcas of the Canadian Arctic (OCA), lancé en 2005, est mené en collaboration avec le milieu universitaire, des gouvernements fédéral, provinciaux et territoriaux, l'industrie, des sociétés d'experts-conseils et des organisations Inuites. Une étude portant sur les épaulards dans l'Arctique canadien a été amorcée en 2006 et s'est poursuivie en 2007 et 2008, en vue de surveiller cette espèce dans cette région.
On recueille les connaissances ancestrales Inuites concernant les épaulards depuis août 2007; les premiers efforts portaient sur la région de la baie d'Hudson. À ce jour, des entrevues ont été réalisées dans cinq collectivités: Repulse Bay, Igloolik, Hall Beach, Rankin Inlet et Arviat. D'autres entrevues sont prévues entre 2008 et 2010. Jusqu'à présent, les entrevues ont livré une myriade de données locales sur les épaulards de l'Arctique, y compris la répartition, la migration et les habitudes migratoires, la saisonnalité et les répercussions de la prédation sur d'autres espèces de mammifères marins.
Simultanément, une base de données sur les observations a été créée et évaluée en vue de déterminer la fiabilité générale des observations, l'accent étant mis sur l'identification des espèces, le type d'observateur et la précision temporelle et spatiale. Une base de données photographiques est également en cours d'élaboration, ce qui permetttra d'identifier les épaulards individuellement.
3.1.2 Le savoir écologique ancestral et la science occidentale
Mike Hammill
On peut définir le savoir écologique ancestral (SEA) comme la somme de connaissances accumulées par des personnes qui, leur vie durant, ont procédé à des observations et acquis de l'expérience, sans le concours de la science occidentale. L'utilisation du SEA et son intégration aux sciences sont demandées par beaucoup car ce savoir fait appel aux ressources et à la longue expérience d'observation de la population locale, et peut fournir des données historiques antérieures aux données scientifiques dans les régions éloignées. En revanche, la science occidentale repose sur une approche expérimentale et la vérification des hypothèses afin d'obtenir de l'information sur des processus naturels.

Béluga marqué
Photo: Véronique Lesage
Le SEA et les sciences diffèrent pour ce qui est de l'information écologique qu'ils livrent, tant au plan de l'intensité observationnelle que du territoire couvert. Ces différences peuvent donner lieu à des conclusions distinctes sur la répartition saisonnière et le regroupement des populations de bélugas et donc influencer les décisions de gestion. À ce jour, peu d'études ont tenté de comparer de manière analytique ces deux ensembles de données. La collecte du SEA dans un format structuré en vue de son examen pour comprendre les points forts et les faiblesses demeure un défi pour les scientifiques et les détenteurs de ce savoir.
Dans la présente étude, l'information sur les mouvements et le regroupement de bélugas a été recueillie dans le cadre d'entretiens avec 427 chasseurs dépositaires du savoir ancestral, ce qui a donné lieu à 3 253 enregistrements saisis dans une base de données par la Makivik Corporation, de Kuujjuaq, au Québec. Des émetteurs satellites ont été installés sur 30 bélugas dans l'est de la baie d'Hudson, au Canada. On a comparé la répartition et les déplacements saisonniers à l'aide d'approches utilisant le SIG qui permettaient un agencement commun des ensembles de données.
Les deux ensembles de données faisaient état de rassemblement estival dans les estuaires (figure 4). Les données de télémesure ont révélé que les bélugas fréquentaient beaucoup la haute mer puisque 76 % des emplacements se trouvaient à plus de 15 km de la côte du Québec. Toutefois, ces déplacements au large ne ressortaient pas des données du SEA, où 83 % des enregistrements indiquaient que les bélugas se limitaient aux régions côtières. Les données de télémesure ont également montré que les bélugas restaient dans la région de l'arc de la baie d'Hudson tout au long de l'été, mais quittaient cette région pour passer l'hiver au large de la côte du Labrador. Les données du SEA ont révélé la présence de bélugas à proximité de la côte du Nunavik pendant l'été, et dans la région de l'arc de la baie d'Hudson en hiver.
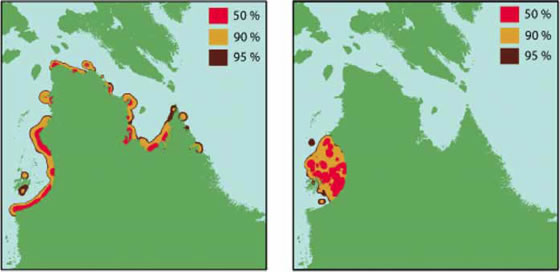
Figure 4. Domaines vitaux en été. Calcul de probabilité de la présence dans le domaine vital de 50, 90 et 95 % d'après les données du SEA, les données de télémesure utilisant les emplacements de 30 baleines. a) SEA pour l'été; b) télémesure pour l'été.
Les résultats des deux méthodes soulignent combien il est important de comprendre la méthode de collecte des données. Les deux méthodes correspondent à différentes méthodes d'échantillonnage qui possèdent leurs avantages et leurs faiblesses. La télémesure par satellite fournit de l'information indépendante sur les déplacements des baleines, en particulier les baleines d'origine connue, dans ce cas, des baleines de la population de l'est de la baie d'Hudson (EBH). Toutefois, relativement peu d'émetteurs ont été installés (n=30) et en moyenne, ils se détachent des animaux après trois ou quatre mois. Les observations des détenteurs du SEA couvrent une plus longue période et une plus grande superficie, mais les observations sont principalement côtières, car il est dangereux de chasser au large dans de petites embarcations. Les données du SEA ne révèlent rien sur l'hivernage au large de la côte du Labrador en raison des difficultés inhérentes aux déplacements en hiver, au raccourcissement des jours et au grand territoire où passe le béluga.
Toutefois, les dépositaires du SEA ont remarqué que certaines baleines passaient l'hiver dans l'EBH. D'où viennent ces baleines? Certaines baleines de la population de l'EBH passent-elles en fait également l'hiver dans cette région? Ou celles qui y hivernent appartiennent-elles à une autre population qui passe l'été ailleurs, dans la baie James par exemple? Les données du SEA indiquent également que l'on trouve des bélugas le long de la côte du Nunavik pendant l'été. Les individus observés dans le détroit d'Hudson l'été sont-ils simplement des individus isolés des populations en migration qui sortent du détroit d'Hudson l'été? Ou appartiennent-ils à d'autres populations reliques qui passent l'été dans cette région?
D'après cette étude, il est évident que les deux ensembles de données peuvent fournir de l'information complémentaire et quand ces données concordent, les conclusions seront probablement plus fiables. Toutefois, quand les différentes méthodes donnent lieu à des conclusions différentes, il importe de comprendre le pourquoi de ces différences. Dans la plupart des cas, il faut bien comprendre les différentes méthodes d'échantillonnage employées, leurs points forts et leurs faiblesses.
3.2 Habitudes
3.2.1 Caractéristiques de plongée et estimations de l'observabilité des baleines boréales dans l'est de l'Arctique d'après la télémesure par satellite
Larry Dueck

Baleine boréale et béluga remontant à la surface dans une polynie
Photo: Steve Ramsay
Le Canada compte deux populations de baleines boréales, l'une qui passe l'été dans la mer de Beaufort dans l'ouest de l'Arctique, et l'autre qui réside dans les eaux de l'est de l'Arctique canadien et dans l'ouest du Groenland. On croyait autrefois que deux populations particulières se retrouvaient dans la population de l'est de l'Arctique. Toutefois, les données que nous possédons nous portent à croire qu'il n'existe en fait qu'une seule et même population, probablement séparée selon la saison en fonction de l'âge et de la reproductivité. La localisation par satellite vient corroborer cette conclusion.
La localisation et la surveillance du comportement de la baleine au moyen d'enregistreurs à liaison satellite archivant les données ont grandement aidé à recueillir des renseignements que l'on n'aurait pu recueillir autrefois en raison du caractère insaisissable de l'espèce qui évolue dans des eaux septentrionales. Le suivi des déplacements de baleines boréales sur de longues distances a montré que cette espèce, dont on pensait qu'elle avait deux aires de distribution distinctes, partage en réalité les aires d'estivage et d'hivernage. L'analyse des déplacements et l'identification du lieu de résidence saisonnière ont livré de nouvelles données sur les voies migratoires, le cycle biologique et l'importance possible d'un habitat particulier.
Grâce à la technologie de pointe de télémesure par satellite, les animaux marqués ont également fourni de l'information sur le comportement de plongée, indiquant que les baleines boréales peuvent descendre à des profondeurs allant jusqu'à 400 m. Au nombre de l'information précieuse fournie par l'examen du comportement de plongée, la proportion du temps passé sous l'eau est importante quand on estime l'abondance de baleines boréales. Il s'avère que les baleines boréales passent près de 75 % de leur temps à des profondeurs supérieures à 4 mètres.
3.2.2 Écologie du phoque annelé
Steve Ferguson

Phoque annelé
Photo: J. Blair Dunn
Les phoques annelés représentent la majeure partie de la chasse de subsistance de mammifères marins pratiquée par les Inuits et ils constituent la principale ressource alimentaire des ours polaires. Les adaptations évolutives des phoques annelés en vue d'exploiter l'habitat de banquise côtière pour la reproduction et la survie pourraient exposer l'espèce à des problèmes très importants par suite du réchauffement planétaire prévu. Des préoccupations sont soulevées quant-à la baisse possible du nombre de phoques annelés dans l'ouest de la baie d'Hudson tel qu'indiqué par les dires des chasseurs, la réduction du taux de grossesse, la baisse de survie des nouveau-nés, la structure d'âges plus vieille et la réduction de la croissance et du nombre d'ours polaires. Quatre relevés aériens qui ont été réalisés entre 1995 et 2000 ont évalué que la taille de la population diminuait, s'établissant à 45 000 phoques en 2000 par rapport à 70 000 en 1995.
Un relevé aérien a été réalisé en mai 2007 et on l'a procédé à la capture et au marquage de phoques annelés en septembre 2007. Le but de ce travail sur le terrain était d'effectuer des prélèvements et de marquer les phoques, afin de mieux comprendre leurs déplacements et leur écologie d'alimentation dans la baie d'Hudson. Les phoques munis d'un enregistreur à liaison satellite fourniront de l'information sur le nombre de fois que les phoques ne sont pas visibles à partir des aéronefs et permettront ainsi de corriger les estimations des relevés aériens. L'information biologique de base, comme l'état et la morphologie, ainsi que des échantillons prélevés de phoques, vivants et morts, ont également été recueillis.
3.2.3 Mise au point de la méthode de surveillance acoustique passive pour surveiller de manière non intrusive les baleines dans leur environnement avec des exemples provenant de la voie navigable du Saint-Laurent et de l'Arctique
Yvan Simard
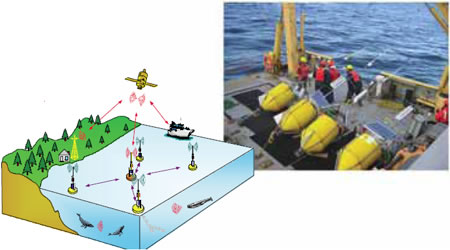
Figure 5. Réseau de bouées intelligentes à liaison radio et satellite WOW pour la localisation en temps réel des baleines grâce à la SAP (photo: Yvan Simard)
Les mammifères marins utilisent de manière intensive l'acoustique sous-marine pour communiquer, naviguer et déceler les proies et les prédateurs. À l'instar des oiseaux, de nombreuses espèces et de nombreux sous-groupes peuvent être identifiés en fonction de leurs cris particuliers. L'enregistrement de ces cris caractéristiques révèle alors la présence de ces espèces dans l'aire de surveillance. Comme un son se propage très efficacement dans l'eau, l'aire de détection peut être vaste, supérieure à 100 kilomètres dans des conditions favorables à des chants à basse fréquence. Ceci dépasse grandement la portée des détections visuelles. Ainsi, les scientifiques spécialistes des mammifères marins évoluant dans diverses disciplines ont travaillé pendant plusieurs décennies afin d'exploiter ce potentiel acoustique pour déceler et surveiller de manière non intrusive les baleines dans leur environnement. Grâce au développement rapide des connaissances et de la technologie dans ce domaine, une nouvelle méthode, appelée surveillance acoustique passive (SAP), voit le jour (figure 5).
On a utilisé divers systèmes de SAP dans le monde, allant des simples systèmes d'enregistrement et à un seul hydrophone jeté par-dessus bord à de vastes systèmes militaires reliés à la côte par câble pour la surveillance acoustique de tous les bassins océaniques. En collaboration avec l'Université du Québec à Rimouski, un programme de recherche a été lancé en 2002 afin d'élaborer et de tester des systèmes de SAP pour la surveillance des baleines dans l'estuaire du Saint-Laurent.
Après avoir mis au point un hydrophone autonome appelé AURAL pour enregistrer des fichiers sonores, on a déployé une série de dispositifs de ce genre dans le parc marin du Saguenay–Saint-Laurent pendant les étés 2003, 2004 et 2005, afin d'examiner différentes configurations possibles pour déceler et localiser le chant du rorqual bleu et du rorqual commun. Plusieurs méthodes de localisation ont été étudiées afin de repérer les baleines dans le bassin de 75 km de longueur; malgré la difficulté particulière de la zone à l'étude, la modélisation de la propagation a été combinée aux caractéristiques des bruits pour montrer que la technologie parvient à déceler et localiser avec une grande efficacité les rorquals bleus et les rorquals communs d'après leurs chants. Une étude est en cours en vue de surveiller la population locale de bélugas.
Le plan pour la prochaine génération de ces systèmes est l'exploitation en temps réel et la détection et localisation automatiques des chants par l'intelligence artificielle. Par ailleurs, un projet de recherche triennal visant à élaborer un réseau de bouées intelligentes complémentaires a été mené à bien. Ce système sera mis à l'essai sur le terrain dans les années à venir. Entre-temps, des hydrophones AURAL ont été déployés en 2004 à plusieurs endroits de l'Arctique et de la baie d'Hudson en vue de surveiller les changements temporels dans l'occupation saisonnière des baleines boréales, des bélugas et des phoques barbus. Les résultats commencent à révéler clairement un lien avec les changements dans l'état des glaces.
3.3 Écologie alimentaire et régime alimentaire
3.3.1 Différences entre les sexes dans l'utilisation de l'habitat, la fréquence de l'alimentation et le régime alimentaire chez les phoques gris
Don Bowen

Phoques gris mâle (arrière-plan) et femelle (avant-plan) adultes illustrant la différence de taille selon le sexe
Photo: Don Bowen
De nombreuses espèces animales se séparent en fonction du sexe. Une telle séparation peut également être sociale de nature, ou écologique, ou les deux. Les phoques gris, comme de nombreux grands mammifères, sont de taille sexuellement dimorphe. Pour étudier la séparation de l'habitat des phoques gris en fonction du sexe, on a recueilli les traces satellites de 95 adultes se reproduisant à l'île de Sable entre 1995 et 2005 (figure 6). Les différences étaient le plus prononcées juste avant (octobre-décembre) et immédiatement après (février-mars) la reproduction. Pendant ces deux périodes, les mâles fréquentaient principalement les aires le long du rebord de la plateforme continentale, tandis que les femelles utilisaient surtout les aires au milieu de la plateforme. Ces différences peuvent servir à maximiser la bonne forme en réduisant la rivalité intersexuelle pendant les principales périodes de prospection alimentaire.
La saison et le sexe expliquent en grande partie la variation observée dans le régime alimentaire des adultes. Les estimations du régime alimentaire ont été établies d'après l'analyse des acides gras (les composantes du gras) des biopsies du petit lard prélevés sur des individus en liberté à l'aide d'une méthode appelée analyse quantitative de la signature des acides gras. Les différences étaient le plus flagrantes pendant la période de prospection alimentaire suivant la reproduction, quand l'absorption d'énergie est importante pour permettre aux femelles d'accumuler des stocks de nutriments qui leur sont utiles en cas de grossesse. Les femelles sélectionnaient moins d'espèces de proie mais de qualité plus élevée au printemps que les mâles. On n'a observé aucune différence entre les sexes dans le régime alimentaire des juvéniles.
On a traditionellement déterminé la fréquence de l'alimentation des phoques d'après l'état de la digestion des proies dans le contenu stomacal. Ces estimations sont plutôt imprécises et ne sont pas forcément représentatives de l'ensemble des déplacements effectués par les animaux en quête de nourriture. Dans une nouvelle étude, de petits émetteurs radios ont été placés dans l'estomac de phoques gris adultes. Ces émetteurs consignaient en continu les changements dans la température de l'estomac associés à l'alimentation. Les résultats ont révélé que le nombre d'événements d'alimentation est généralement plus élevé chez les mâles que chez les femelles, tout comme le temps consacré à l'alimentation chaque jour. Les phoques se nourrissaient en moyenne 57,8 % de la journée et avaient en moyenne 1,7 repas par jour. Les phoques gris avaient tendance à avoir de nombreux événements d'alimentation distincts, de longues périodes séparant chaque événement, comme on s'attend d'un grand carnivore. Ces résultats fournissent de nouvelles données sur le fondement des différences entre les sexes dans la plongée et le régime alimentaire.
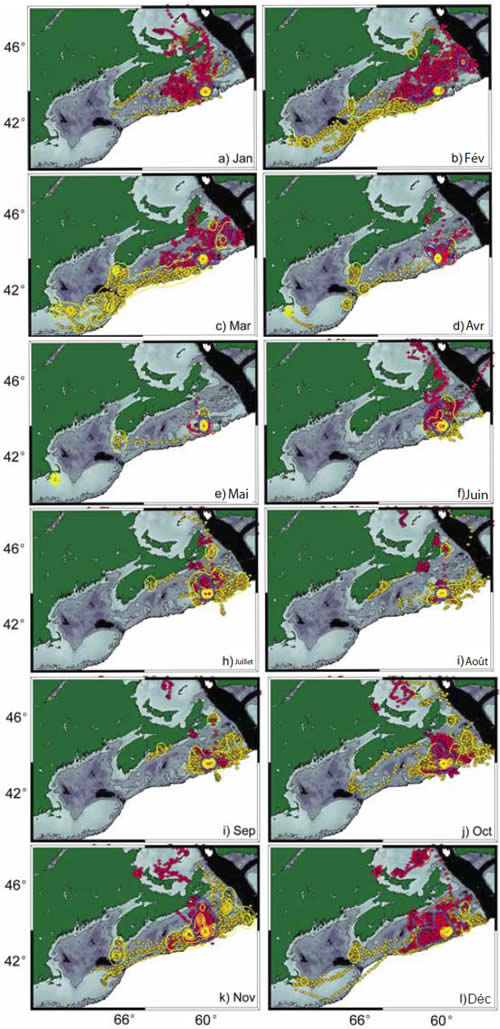
Figure 6. Répartition mensuelle des phoques gris adultes mâles (jaune) et femelles (rouge) selon la localisation satellite d'Argos. Les lignes pleines entourant les secteurs correspondent aux densités des noyaux indiquant les aires de forte utilisation.
3.3.2 Régime alimentaire du phoque du Groenland et du phoque à capuchon
Garry Stenson

Phoque du Groenland femelle et son petit
Photo: DFO

Famille de phoques à capuchon
Photo: Mike Hammill
Les phoques du Groenland et les phoques à capuchon sont deux des mammifères marins les plus abondants dans l'Atlantique Nord et jouent par conséquent un rôle important dans la structuration de l'écosystème. D'importantes recherches ont été réalisées au cours de la dernière décennie pour déterminer leur rôle écologique et l'incidence possible de la prédation du phoque sur la dynamique de la population de leurs proies. Toutefois, il s'agit de questions extrêmement complexes pour lesquelles il n'existe pas de réponses toutes faites.
Le développement de la télémesure par satellite a donné aux scientifiques la possibilité de mieux comprendre les déplacements et l'utilisation de l'habitat des phoques en liberté. Les phoques du Groenland et les phoques à capuchon sont des espèces pélagiques qui passent une grande partie de leur temps dans l'eau. Les phoques du Groenland se trouvent principalement le long des plateformes continentales où ils descendent à des profondeurs relativement peu élevées (100-200 m), même si on en a observé qui plongeaient à une profondeur pouvant atteindre 800 m. Par contre, les phoques à capuchon habitent les rebords des plateformes continentales et les pentes en eaux profondes. Les phoques à capuchon plongent régulièrement à des profondeurs supérieures à 300 m, et à l'occasion, à plus de 1 500 m.
Les deux espèces se nourrissent de divers poissons et invertébrés. Le régime alimentaire exact varie selon l'âge, le sexe, l'emplacement, la saison et l'année. En général, les phoques du Groenland se nourrissent d'une variété de petits poissons fourrages, comme le capelan, la morue arctique (morue polaire en Europe, Boreogadus saida), le hareng et le lançon. Leurs proies sont également des invertébrés comme des amphipodes et des crevettes. Même si les phoques à capuchon se nourrissent de nombre de ces espèces, ils ont tendance à consommer de plus grosses quantités d'espèces d'eau profonde comme le flétan noir et le sébaste que l'on trouve le long des rebords de la plateforme.
Traditionnellement, on établissait le régime alimentaire au moyen des morceaux durs trouvés dans les estomacs. Toutefois, chaque méthode présente des biais éventuels susceptibles d'affecter l'exactitude des estimations du régime alimentaire. De nouvelles techniques comme l'analyse de la signature des acides gras et l'analyse d'ADN du contenu stomacal fournissent de nouvelles données sur les régimes alimentaires à des échelles temporelles et spatiales plus grandes. Elles fournissent également de nouvelles données sur l'importance d'espèces-proies consommées par le phoque du Groenland et le phoque à capuchon et permettent de déterminer l'ampleur des biais associés à chaque méthode.
La consommation d'importantes espèces-proies par les phoques dans l'Atlantique canadien a été évaluée à l'aide de modèles bioénergétiques. Les phoques du Groenland sont d'importants prédateurs au large de la côte est de Terre-Neuve et dans le nord du golfe du Saint-Laurent, tandis que les phoques à capuchon se nourrissent principalement au large de Terre-Neuve et aux alentours du Bonnet flamand.
Plusieurs études ont tenté de déterminer l'incidence des phoques sur les stocks de poissons dans le nord-ouest de l'Atlantique, en particulier l'incidence des phoques du Groenland et des phoques gris sur la morue. Il appert généralement que même si les phoques consomment de grandes quantités d'espèces de poisson commerciales et d'importantes espèces fourrages, l'incidence de ces prélèvements sur les stocks actuels de poissons est difficile à déterminer. Les phoques sont d'importants prédateurs de la grande morue et de la jeune morue et pourraient bien jouer un rôle dans le non-rétablissement des stocks de morues, mais on ne saurait leur imputer une part importante de la mortalité dans la plupart des régions, et par conséquent, l'incidence totale de leur prédation ne peut être déterminée. Souvent, les estimations de la consommation de morues en fonction de leur âge par les phoques sont en contradiction avec la mortalité élevée observée parmi les groupes plus âgés. On ignore pratiquement tout de la réponse fonctionnelle des phoques aux changements dans l'abondance des proies, des autres sources de mortalité ou des effets écosystémiques possibles comme la compétition pour la consommation de poisson fourrage et la rétroaction positive de la prédation du phoque sur les poissons piscivores.
3.4 Interactions entre le prédateur et la proie
3.4.1 Rapport de l'Atelier national sur les répercussions des phoques sur les populations de poissons dans l'est du Canada
Don Bowen

Morue capturée dans un chalut
Photo: DFO
Une réunion de cinq jours s'est tenue en novembre 2008 à Halifax, en Nouvelle-Écosse et portait sur des sujets se rapportant aux répercussions éventuelles des phoques sur les stocks de poissons dans l'est du Canada. On formule pour hypothèse que les phoques ont cinq effets naturels négatifs sur les populations de proies :
- prédation;
- compétition;
- transmission de parasites causant une mortalité accrue des poissons;
- perturbation du frai, ce qui entraîne une baisse du succès de reproduction; et
- autres effets indirects sur le comportement du poisson causés par le risque de prédation du phoque. Il s'agit du deuxième de deux ateliers et exposés axés sur les nouvelles analyses et les résultats du modèle découlant des travaux dont il a été question au premier atelier. Trente participants provenant du Canada, de la Norvège et des États-Unis ont pris part à la deuxième réunion. Des membres de l'industrie de la pêche, des étudiants des cycles supérieurs de l'Université Dalhousie et des scientifiques intéressés de l'Institut océanographique de Bedford ont également assisté à des séances de la réunion. Le principal objectif de ces ateliers était d'examiner ce qui est connu, de relever les lacunes dans nos connaissances et de formuler des conclusions concernant les répercussions du phoque sur les stocks de poissons dans l'est du Canada. Un rapport de la réunion sera établi au début de 2009.
3.4.2 L'importance de la prédation du phoque gris sur la morue
Don Bowen

Diverses catégories de taille de la morue
Photo: Tom Hurlbut
L'écosystème de la plateforme Scotian a subi d'importants changements. Les prédateurs du sommet de la chaîne autrefois abondants ne représentent plus qu'une petite fraction de leur nombre historique, et une grande partie de la structure communautaire actuelle est maintenant dominée par les poissons pélagiques et les invertébrés. Dans ce réseau trophique, les populations de morue et de phoque gris ont affiché des tendances inverses. Depuis 1984, les populations de morue ont diminué de manière exponentielle à un taux moyen de 17 % par an. Parallèlement, les phoques gris ont continué d'augmenter de manière exponentielle, à un taux s'élevant en moyenne à 12 %.
L'incidence des phoques gris sur la dynamique de la morue a été réexaminée à la lumière de plus de 30 années de données sur les tendances démographiques de la morue et du phoque gris, en tenant compte de nouvelles données sur le régime alimentaire du phoque et la répartition saisonnière. La fermeture de la pêche à la morue il y a plus de 10 ans a permis de mieux évaluer les taux de mortalité naturelle.
Entre 1993 et 2000, la morue représentait en moyenne plus ou moins 2 % du régime alimentaire du phoque gris. Depuis la fermeture de la pêche, les phoques gris sont à l'origine d'un niveau élevé de mortalité instantanée, soit 21 % de la mortalité totale en moyenne. Toutefois, la mortalité naturelle de la morue pendant cette période est en grande partie imputable à des sources inconnues. Néanmoins, comme l'abondance de la morue a diminué pendant l'étude, la consommation de morue par le phoque gris a bel et bien contribué à l'échec du rétablissement des stocks de morue.
3.4.3 Stratégies antiprédatrices des cétacés à fanons
John Ford

Le rorqual commun peut nager à une vitesse de plus de 30 km/h pour échapper aux épaulards qui le poursuivent.
Photo: Mark Malleson
On a émis récemment l'hypothèse selon laquelle les épaulards – le prédateur aquatique le plus important de la plupart des espèces de mammifères marins – étaient à l'origine de l'effondrement des populations de phoques, de lions de mer et de loutres de mer dans le Pacifique Nord. Selon cette hypothèse, la chasse industrielle de grands cétacés à fanons au XXe siècle aurait éliminé cette source de proie préférée des épaulards chasseurs de mammifères et les aurait contraints d'opter pour d'autres espèces-proies. Cette idée est loin de faire l'unanimité pour diverses raisons, entre autres le fait que rien ne nous indique que l'épaulard a jamais été un important prédateur des grands cétacés à fanons. Toutefois, ce point soulève une question intéressante – pourquoi les épaulards n'ont-ils pas pour proie régulière les grandes baleines?
Pour mieux comprendre la tactique et les stratégies que les cétacés à fanons utilisent pour éviter ces prédateurs, on a compilé et examiné des rapports publiés et des observations non publiées des interactions prédateur-proie visant l'épaulard et le cétacé à fanons. Cette étude a révélé que les cétacés à fanons adoptaient deux stratégies antiprédatrices distinctes et opposées lorsqu'ils sont harcelés ou attaqués par des épaulards: la stratégie de combat, où les baleines se regroupent et se défendent ensemble, et la stratégie de fuite, où les baleines s'enfuient rapidement et, si elles se font rattraper et attaquer physiquement, n'essaient pas de se défendre.
Au nombre des espèces de combat, mentionnons les baleines boréales, les baleines noires, les rorquals à bosse et les baleines grises, qui ont tous des formes de corps relativement robustes et massives et sont des nageurs relativement lents mais agiles. En général, ces baleines échappent aux attaques des épaulards en roulant sur elles-mêmes, en faisant des éclaboussures à la surface de l'eau et en attaquant leurs prédateurs avec la pointe de leur queue. Par contre, les espèces qui s'échappent – le petit rorqual, le rorqual boréal, le rorqual commun et le rorqual bleu – ont tous la même forme de corps hautement aérodynamique et nagent vite. Ces baleines arrivent généralement à échapper aux épaulards qui les poursuivent en s'enfuyant à des vitesses élevées allant de 20 à 40 km/h.
De nombreux aspects du cycle biologique, du comportement et de la morphologie des cétacés à fanons coïncident avec leur stratégie antiprédatrice, et il semble que l'évolution de ces traits ait été façonnée par la sélection en vue d'une prédation réduite. Par exemple, les espèces qui se battent se rassemblent souvent pour la reproduction dans des eaux côtières peu profondes, où elles sont mieux en mesure de se défendre et de défendre leurs petits contre les épaulards. La plupart des espèces qui s'échappent pénètrent rarement dans des eaux restreintes qui pourraient entraver leur fuite. Les espèces qui se battent ont également tendance à avoir sur le corps des incrustations (callosités et bernacles) qui peuvent servir d'armure ou d'armes en vue d'une meilleure défense.
En gros, il semble que ces stratégies antiprédatrices divergentes adoptées par les cétacés à fanons permettent d'éviter la prédation par les épaulards, en particulier les adultes. Par conséquent, les épaulards attaquent rarement ces grandes baleines et ciblent plutôt les mammifères plus petits, plus faciles à attraper, qui sont des proies plus profitables. Il semble douteux que la réduction du nombre de grandes baleines par suite de la chasse ait eu une grande incidence sur les habitudes alimentaires des épaulards.
3.5 Habitat des mammifères marins
3.5.1 Les phoques communs à Terre-Neuve et au Labrador: nouvelles données
Becky Sjare

Phoque commun au repos
Photo: DFO Quebec
Le phoque commun est l'une des sept espèces de pinnipèdes que l'on trouve dans le nord-ouest de l'Atlantique. Même si cette espèce est bien étudiée dans une grande partie de son aire de répartition, on en sait relativement peu à propos de l'écologie et du statut démographique de ces phoques dans les eaux de Terre-Neuve et du Labrador. Les objectifs de ce projet étaient de déterminer les profils des contaminants de base pour les phoques communs et leurs principales espèces-proies, de recueillir les données écologiques nécessaires afin d'évaluer si l'espèce pourrait être un bon indicateur de la santé de l'écosystème et de commencer à envisager les façons d'utiliser les données pour la surveillance des effets cumulatifs à long terme des aménagements côtiers futurs se rapportant à la production pétrolière en mer.
Selon un nombre limité de relevés de reconnaissance par bateau, des dénombrements opportunistes des individus montant sur l'échouerie effectués à partir de la côte et des entretiens avec des pêcheurs, les phoques communs sont relativement répandus mais leur répartition est clairsemée, et leur densité faible. Les exceptions sont les côtes nord-ouest et nord-est de Terre-Neuve et du Labrador, où l'on a fait peu d'observations. Il semble que l'abondance locale de phoques à des échoueries connues le long des côtes sud et sud-ouest a augmenté; l'abondance sur des sites situés plus au nord demeure faible.
En général, les phoques communs sont connus pour consommer une grande variété de proies et leur régime alimentaire varie selon la région, la saison et l'âge. Les analyses des échantillons du contenu stomacal prélevés entre 1985 et 2003 ont révélé la consommation d'une trentaine d'espèces différentes de poissons et d'invertébrés, mais dix espèces représentaient près de 95 % du contenu. La plie rouge, la morue arctique, le chaboisseau à épines courtes et la morue franche étaient les plus importantes.
Entre 2001 et 2003, un total de 66 échantillons de tissus ont été prélevés dans toute la province et analysés en vue de déceler les niveaux de mercure total, d'éléments traces et de polluants organiques persistants (POP). Les concentrations d'éléments traces et de mercure total étaient généralement comparables à celles présentes dans d'autres espèces de phoques du nord. Le mercure total, le sélénium et le cadmium ont affiché la plus grande variabilité entre les sites d'échantillonnage et dans ces sites; les phoques sur lesquels des échantillons ont été prélevés le long des côtes sud et est de la province avaient les concentrations les plus élevées de cadmium rénal. La source du cadmium n'est pas encore connue.
Selon la série de POP examinés, les phoques communs des eaux terre-neuviennes sur lesquels on a effectué des prélèvements étaient moins contaminés que ceux provenant de la population de l'estuaire du Saint-Laurent, mais leur contamination était généralement similaire à ceux du sud du golfe du Saint-Laurent. Les concentrations de mirex et de BPC étaient de 5 à 10 fois supérieures dans la population de l'estuaire tandis que celles de DDT et de chlordane étaient de deux à cinq fois plus élevées que chez les phoques de Terre-Neuve. Les niveaux de POP chez les mâles matures étaient plus élevés que chez les femelles, mais il n'existait aucune différence entre les sexes des juvéniles.
Ces nouvelles données sur les phoques communs jetteront les bases des études écologiques ultérieures, des évaluations des populations, de l'établissement d'indicateurs écosystémiques pour les régions côtières et de la recherche portant sur le mode d'accumulation des contaminants dans les chaînes trophiques côtières.
3.5.2 Caractéristiques sexospécifiques des troupeaux de loutres de mer de même sexe
Linda Nichol

Loutre de mer
Photo: Brian Gisborne
Les regroupements de loutres de mer de même sexe peuvent inclure jusqu'à 100 individus se reposant ensemble dans l'eau. Les mâles et les femelles se séparent et occupent spatialement des aires de regroupement distinctes. Ces regroupements d'individus de même sexe ont généralement lieu au même endroit pendant plusieurs années. D'après les observations sur le terrain, il semble y avoir des différences dans les caractéristiques des lieux de regroupement utilisés par les mâles et les femelles ainsi que des différences dans la formation quotidienne de regroupements. Les regroupements de femelles sont plus nombreux que ceux de mâles.
Dans cette étude, les caractéristiques de 49 sites de regroupement ont été mesurées dans un environnement de SIG. Les caractéristiques physiques incluent la profondeur moyenne, la distance par rapport aux grandes et petites masses continentales émergées, l'exposition et la complexité du fond. Les regroupements de mâles étaient en moyenne mieux protégés au sud-est que les regroupements de femelles. Plusieurs regroupements de mâles se trouvaient dans des bras, contrairement à ceux de femelles. Les regroupements de mâles se trouvaient plus près des grandes masses continentales émergées et ceux des femelles, plus près des petites masses continentales émergées. Les regroupements des deux sexes ont été observés à des profondeurs inférieures à 20 mètres.
Les différences sexospécifiques dans les caractéristiques des lieux de regroupement reflètent probablement les différences dans les besoins et les contraintes avec lesquels doivent composer les loutres de mer mâles et femelles. Les femelles adultes donnent naissance à un seul petit chaque année et s'en occupent pendant six à huit mois. Les grandes masses continentales émergées constituent des sources possibles de prédateurs terrestres et de pygargues à tête blanche, connus pour s'attaquer aux petits. Les terres émergées de moins de 50 mètres étaient généralement des écueils offrant un habitat peu profond variable, caractéristiques propres à diverses aires d'alimentation en eaux peu profondes. Pour trouver les invertébrés benthiques dont se nourrissent les loutres de mer, les femelles doivent laisser leur petit sans surveillance à la surface pendant qu'elles plongent. Il est possible que les femelles ayant des petits préfèrent les faibles profondeurs.
3.5.3 De la physique aux baleines: l'exemple de l'aire d'alimentation des cétacés à fanons de l'estuaire du Saint-Laurent
Yvan Simard
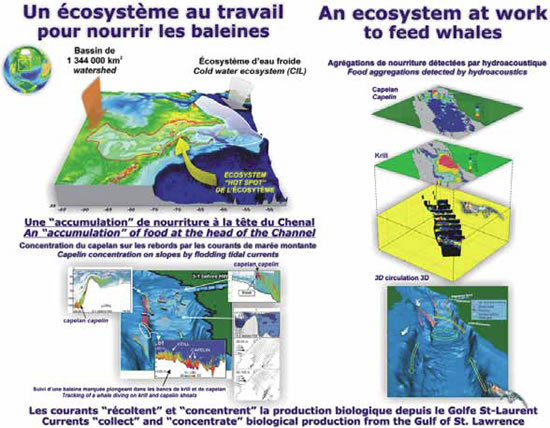
Figure 7

Rorqual commun
Photo: John Ford
Les cétacés à fanons du nord-ouest de l'Atlantique migrent chaque année dans le golfe du Saint-Laurent pendant la saison sans glace pour reconstituer leurs réserves de gras en s'alimentant de leurs proies préférées. Cette alimentation annuelle intensive est essentielle à la santé des individus, à leur reproduction et à la croissance de la population. La question est la suivante: qu'est-ce qui occasionne et contrôle ces regroupements?
Un plan de recherche systématique a utilisé une approche écosystémique pour répondre à cette question et mettre en lumière les processus océanographiques déterminant les regroupements de proies dans une aire d'alimentation traditionnelle des baleines dans le bassin hydrographique du Saint-Laurent. Plusieurs espèces, entre autres le rorqual bleu, le rorqual commun et le petit rorqual, visitent régulièrement cette aire, située à l'extrémité amont de leur route migratoire annuelle. Ce retour continu témoigne de l'existence d'un processus pérenne qui maintient constamment les proies regroupées à la tête du chenal.
Le programme de recherche a montré que cette aire était le site du regroupement le plus abondant de krills dans le nord-ouest de l'Atlantique. Le mécanisme (figure 7) responsable est le drainage du krill adulte d'une grande partie du golfe du Saint-Laurent. Les interactions tidales des courants et du krill avec les pentes du chenal entraînent une concentration du krill adulte dans le courant ascendant. Ce courant favorise l'accumulation de krills dans le parc marin. Des espèces de poissons fourrages pélagiques comme le capelan se réunissent aussi dans cette aire. D'autres recherches sur la dynamique de regroupement à la tête du chenal et à l'entrée du fjord du Saguenay sont en cours.
Des mesures hydro-acoustiques intensives de la répartition du krill dans l'ensemble du golfe du Saint-Laurent étaient requises pour brosser un tableau complet de l'aire d'alimentation permanente exceptionnelle des cétacés à fanons dans l'est du Canada. La modélisation a également fait ressortir d'autres zones de regroupement du krill dans le bassin hydrographique du Saint-Laurent, qui coïncident avec les données publiées provenant d'un relevé aérien du golfe.
4.0 Répercussions anthropiques sur les mammifères marins
4.1 Chasses commerciales et chasses de subsistance
4.1.1 Baleines boréales dans l'est de l'Arctique canadien et dans l'ouest du Groenland
Jeff Higdon

Chasse fructueuse à la baleine boréale
Photo: Jack Orr
La chasse commerciale des baleines boréales dans l'est du Canada et dans l'ouest du Groenland remonte à 1530, quand des baleiniers basques les ont chassées dans le détroit de Belle-Isle. Les Inuits du Canada et de l'ouest du Groenland ont également chassé la baleine boréale à des fins de subsistance et de commerce pendant des siècles. La planification du rétablissement des baleines boréales requiert une estimation de la taille de la population avant leur exploitation afin d'établir des objectifs de rétablissement. On peut estimer cette population en utilisant un modèle démographique et une série sur les prélèvements. L'établissement d'une série sur les captures, comme on le mentionne ici, a été la première étape.
La chasse commerciale à la baleine boréale a commencé au début des années 1500 dans la région du détroit de Belle-Isle, et les baleiniers danois et allemands voyageaient jusqu'au détroit de Davis vers la fin des années 1600. La colonisation de l'ouest du Groenland par les Danois et les Norvégiens a commencé en 1721 et ces colons participaient aussi activement à la chasse à la baleine boréale. Les baleiniers britanniques se trouvaient dans le détroit de Davis vers le milieu des années 1700 et se sont rendus dans la baie de Baffin au début des années 1800, donnant une nouvelle impulsion à cette chasse au large de l'île de Baffin, dans le détroit de Lancaster, dans l'inlet Prince-Regent et dans le golfe de Boothia. Les baleiniers américains ont été actifs dans le détroit de Davis dans les années 1700 et à nouveau dans les années 1800, et dans la baie d'Hudson après 1860, où les baleiniers écossais se sont joints à eux. La dernière chasse commerciale à la baleine boréale dans l'Arctique canadien a eu lieu en 1915, quand la population a atteint des effectifs extrêmement faibles et que les expéditions n'étaient plus rentables. On a estimé à 57 507-68 736 baleines le total (minimal) des prélèvements commerciaux entre 1530 et 1915, les plus grosses prises étant réalisées par les Basques et les Britanniques.
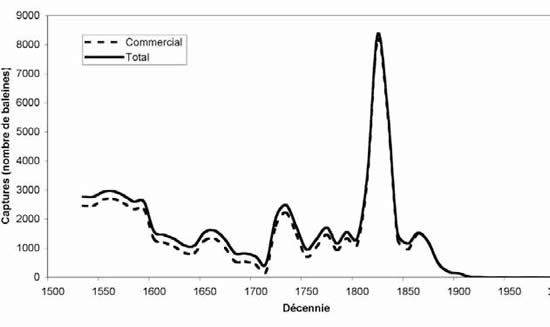
Figure 8. Total estimatif des prélèvements de baleines boréales par décennie dans l'est du Canada et l'ouest du Groenland entre 1530 et 2005, illustrant le faible volume des prises de subsistance des Inuits par rapport aux prises commerciales arvest per decade in eastern Canada and West Greenland from 1530-2005, illustrating how small Inuit subsistence harvests were in comparison to commercial harvests.
La première culture indigène à donner naissance à des chasseurs actifs de la baleine boréale était la culture de Thulé. Après 1500, la chasse à la baleine boréale a diminué, peut-être en raison de conditions climatiques changeantes, d'une raréfaction des baleines attribuable à la chasse pratiquée par les Basques ou d'une combinaison de ces deux facteurs. Les prises estimatives totales entre 1530 et la fin de la chasse commerciale à la baleine s'élèvent à 8 460 baleines. La chasse à la baleine pratiquée par les Inuits a diminué elle aussi après que les baleiniers commerciaux ont surexploité la population de baleines boréales, et on estime que seulement 56 baleines ont été chassées après 1918. L'estimation des prises des Inuits repose sur des données éparses et plusieurs hypothèses; il semble qu'au moins plusieurs parties de la série de données sur les prises soient des sous-estimations. La longue tradition de la chasse à la baleine boréale par les Inuits a subi les effets négatifs de la chasse commerciale. Il existe actuellement une chasse limitée au Nunavut et des chasses similaires prévues au Nunavik et dans l'ouest du Groenland.
Si l'on combine les prélèvements commerciaux inuits après 1530 (sans tenir compte des baleines abattues et perdues et en sachant que les données sont incomplètes pour certaines nations et certaines époques), on évalue à plus de 70 000 baleines le nombre de baleines tuées. Presque toutes (88 %) ont été capturées par des baleiniers commerciaux. Bien que la série soit encore incomplète, elle est plus détaillée que les données disponibles par le passé et devrait améliorer les efforts de modélisation visant à estimer la taille de la population avant la chasse à la baleine (figure 8). Toutefois, toute modélisation devrait évaluer rigoureusement la sensibilité des résultats aux aspects variables de la série sur les prélèvements en raison du nombre d'hypothèses incluses. La qualité des données sur les prises varie considérablement d'une nation et d'une époque à l'autre. Les études de modélisation de la population devront explicitement tenir compte de cette variabilité dans la qualité des données.
4.2 ProvidingFormulation d'avis scientifiques sur les TAC
4.2.1 Gestion du phoque du Groenland, du phoque à capuchon et du phoque gris au Canada
Garry Stenson
Depuis le début des années 1980, le gouvernement canadien utilise un point de référence appelé « taux de remplacement » dans la gestion du phoque du Groenland, du phoque à capuchon et du phoque gris dans l'Atlantique Nord-Ouest. Le taux de remplacement correspond au nombre d'animaux qui peuvent être capturés pendant une année donnée sans entraîner de réduction de la population totale l'année suivante. En 2003, toutefois, le Canada a adopté une nouvelle méthode de gestion des phoques dans le Canada atlantique. Cette méthode, appelée gestion des pêches par objectifs, comprend une approche de précaution, qui privilégie la prudence quand l'information est moins certaine, n'accepte pas l'absence d'information comme motif pour ne pas mettre en œuvre des mesures de conservation et définit, en avance, les règles de décision pour la gestion des stocks quand la ressource atteint des points de référence clairement énoncés. Ces points ou niveaux sont appelés seuils de référence cibles, de précaution (ou tampons) et pour la conservation (limitée ou critique).
L'un des principes de base de l'approche de précaution est la nécessité de tenir compte de l'incertitude associée aux estimations et d'élaborer un fondement pour la prise de mesures même si on n'en sait pas assez sur l'état du stock. Par conséquent, nous établissons une distinction entre les espèces pour lesquelles on dispose d'une quantité considérable de données (espèces bien documentées) et les situations où l'information sur les ressources est plus limitée (espèces peu documentées). Pour les espèces bien documentées, il faut établir au moins trois estimations de l'abondance sur une période de 15 ans, la dernière d'entre elles étant obtenue au cours des cinq dernières années, et disposer d'information à jour (moins de cinq ans) sur la fécondité et la mortalité. Si on ne dispose pas de ces données, les espèces seront jugées mal documentées.
Pour les espèces bien documentées, le seuil de référence pour la conservation (Ncritique) représente l'abondance numérique (estimative) à laquelle les prélèvements continus sont jugés avoir des effets préjudiciables graves et irréversibles sur la population. Toutefois, les estimations de l'abondance sont associées à une grande incertitude et cette incertitude s'accroît à mesure que la projection embrasse un avenir plus lointain. La gestion d'une population près du seuil de référence pour la conservation pourrait accroître la probabilité que la population diminue de manière non délibérée sous le Ncritique. Par conséquent, un seuil de référence de précaution (Ntampon) détermine une fourchette de population « à risque » au sein de laquelle des règles de contrôle de gestion prudente s'appliquent. Quand une population se trouve en deçà du seuil de référence de précaution, les gestionnaires peuvent établir un seuil de référence cible reposant sur des facteurs comme les répercussions sur l'écosystème et les retombées socio-économiques. Tant que la population demeure au-dessus du Ntampon, des stratégies de capture à risque plus élevé peuvent être adoptées. En fait, au Canada, il faut que la probabilité qu'une population se trouve au-dessus du Ntampon soit de 80 % pour que cette population soit estimée en bonne santé.
Si la population se trouve sous le Ntampon (mais au-dessus du Ncritique), les préoccupations en matière de conservation peuvent devenir une priorité et des règles préétablies de contrôle de l'exploitation sont alors appliquées dans le but de rétablir la population afin qu'elle retrouve un niveau supérieur au Ntampon dans un certain nombre d'années. Bien que l'exploitation et d'autres prélèvements par l'homme puissent se poursuivre, les stratégies de gestion exigent une probabilité élevée d'augmentation de la population (ou au contraire, un risque moindre de décroissance).
Une population inférieure au Ncritique est préoccupante sur le plan de la conservation et la poursuite des prélèvements présente un risque inacceptable d'effets préjudiciables graves ou irréversibles. Par conséquent, les mesures de gestion devraient veiller à ce que toute mortalité causée par l'homme soit éliminée.

Figure 9. Méthode de gestion des pêches par objectifs adoptée par le Canada en 2003 avec des seuils de référence établis pour les phoques du Groenland dans le nord-ouest de l'Atlantique.
Actuellement, nous avons établi que le Ncritique correspondait à 30 % du niveau de population le plus élevé estimé ou obtenu par déduction (N30), tandis que le Ntampon correspond à 70 % de ce maximum (N70) (figure 9). À des fins de gestion, nous avons également établi un niveau à 50 %, où des mesures de gestion plus rigoureuses s'imposent. Le seuil de référence de précaution est établi à un niveau assez élevé en raison des caractéristiques de la façon dont les phoques sont évalués et de la structure par âge des prises actuelles. Comme la principale méthode d'évaluation est d'estimer la production de nouveau-nés tous les quatre ou cinq ans et sachant que l'on trouve un nombre élevé de jeunes de l'année dans les prises, on ne peut déterminer l'incidence des captures actuelles avant que les animaux survivants ne se soient reproduits au moins cinq années plus tard. Par ailleurs, les conditions environnementales récentes ont été médiocres et continueront probablement à empirer, dans l'ensemble. Enfin, l'histoire a montré que les populations de mammifères marins augmentent de manière relativement lente, mais peuvent décroître rapidement (p. ex., béluga du Saint-Laurent, rorqual bleu) et que le fait de ne pas tenir compte du facteur d'incertitude peut entraîner de graves dommages (p. ex., morue franche, saumon, oreille de mer).
Le 7 octobre 2007, l'Organisation maritime internationale (OMI) a adopté une proposition (présentée par Transports Canada) en vue de soustraire à la navigation 1 780 kilomètres carrés du bassin Roseway, en les désignant « zones à éviter » par les navires de 300 tonnes de jauge ou plus, seulement en transit, entre le 1er juin et le 31 décembre. La date d'entrée en vigueur est le 1er mai 2008.
4.2.2 Mortalité causée par la chasse chez les phoques à capuchon de l’Atlantique Nord-Ouest
Garry Stenson
Pour déterminer avec précision l’abondance des populations sauvages, il est nécessaire d’obtenir les données les plus complètes possibles sur le niveau de mortalité causée par l’homme. Ceci est particulièrement important pour les espèces exploitées à des fins commerciales ou de subsistance. Dans l’Atlantique Nord-Ouest, les phoques à capuchon, comme les phoques du Groenland, sont tués dans le cadre de la chasse de subsistance au Groenland et de la chasse commerciale dans les eaux méridionales du Canada. Toutefois, contrairement aux phoques du Groenland, un nombre extrêmement faible de phoques à capuchon est prélevé dans l’Arctique canadien ou est prélevé en tant que prises accessoires dans le cadre des pêches commerciales
L’objectif de cette étude était de résumer les données disponibles sur les prélèvements de phoques à capuchon déclarés et d’estimer les taux de mortalité totaux attribuables à la chasse chez cette espèce entre 1954 et 2006. Avant l’imposition des quotas en 1974, les prélèvements canadiens étaient très variables, allant de quelques centaines d’individus à plus de 25 000. Entre 1974 et 1982, les prélèvements ont atteint une moyenne de 12 500 phoques. Toutefois, en raison d’un effondrement des principaux marchés, les prélèvements ont chuté après 1982. Les prélèvements annuels ont été pour la plupart de l’ordre de quelques centaines d’animaux jusqu’au milieu des années 1990. En 1996, un grand nombre de dos bleus ont été capturés illégalement, tandis qu’une subvention pour la viande a entraîné des captures de plus de 7 000 animaux en 1997 et de 10 000 animaux en 1998. Depuis, les prélèvements annuels de phoques à capuchon dans les eaux canadiennes sont demeurés faibles.
Entre 1945 et 1960, des phoques à capuchon ont été chassés par des navires norvégiens. Les prélèvements annuels ont varié de 1 500 à 48 000 individus et ont atteint une moyenne de 14 500 phoques. Entre 1970 et 1978, moins de 1 000 phoques ont été capturés par deux ans par des scientifiques norvégiens. Entre le milieu des années 1950 et 1972, les prélèvements effectués au Groenland se sont situés entre 1 000 et 2 000 individus. Depuis, les captures sont relativement stables, s’établissant en moyenne entre 6 000 et 7 000 phoques.
Entre 1954 et 1960, les prélèvements moyens ont été de 34 800 phoques. De 1974 à 1982, les prélèvements de phoques à capuchon s’élevaient en moyenne à 24 600 individus. La majorité des phoques à capuchon ont été capturés au Groenland. À l’exception de la période s’étendant de 1996 à 1998, durant laquelle les prélèvements canadiens représentaient plus de 70 % des captures annuelles, les prélèvements de phoques à capuchon par le Groenland représentaient plus de 65 % des phoques capturés. Environ 30 % des phoques à capuchon tués sont des jeunes de l’année.
4.2.3 Détermination du total autorisé des captures de bélugas et de narvals du Nunavut
Pierre Richard
La Gestion des pêches et de l'aquaculture (GPA) a demandé au Secteur des sciences de formuler des avis sur le total autorisé de captures visant tous les stocks de bélugas et de narvals du Nunavut. La GPA a également demandé que les avis soient formulés selon son cadre provisoire d'approche de précaution. À cette fin, il faut disposer d'une série d'évaluations sur une période suffisamment longue de la taille de la population, de la mortalité causée par la chasse et des paramètres concurrents de la dynamique de la population. Ceci n'est pas possible pour les espèces peu documentées – les stocks pour lesquels on ne peut estimer les paramètres de la dynamique de la population; la méthode prudente de prélèvement biologique potentiel (PBP) a été utilisée pour déterminer le total autorisé des captures de ces stocks.

Bélugas à la surface de l’eau dans une polynie
Photo: M. Ramsay

Groupe de narvals
Photo: Larry Dueck
On a également comparé les résultats du prélèvement biologique potentiel à un modèle de croissance simple utilisé pour déterminer la probabilité de risque de différents niveaux de déclin. Selon ce modèle, les répartitions de l'incertitude servent à modéliser l'imprécision de la taille de la population et du taux de perte par la chasse de même que l'incertitude associée au taux de croissance de la population. Si l'on compare le modèle de risque au modèle de croissance simple, le total autorisé recommandé des captures débarquées, c'est-à-dire le PBP moins les pertes par la chasse, a une faible probabilité d'induire une tendance à la baisse. À l'avenir, d'autres relevés de ces stocks pourront aider à évaluer la tendance et les paramètres de la dynamique de la population pour ces stocks.
4.3 Prises accessoires de mammifères marins
4.3.1 Prises accessoires de phoques du Groenland dans la pêche de la lompe à Terre-Neuve
Becky Sjare

Prises accessoires de phoques du Groenland dans un casier à crabes
Photo: DFO Newfoundland
espèces de phoques présentes dans les eaux terre-neuviennes se retrouvent prises fortuitement dans des engins de pêche actifs, comme des chaluts, et des engins passifs, comme des filets maillants. À Terre-Neuve, on retrouve des phoques du Groenland dans la majorité des pièges, principalement dans les filets maillants installés pour la lompe.
On trouve la lompe dans toutes les zones côtières de la province et de grandes concentrations sur le Banc de Saint-Pierre. La pêche utilise des filet maillants à filament simple dont la taille des mailles varie entre 25 et 27 cm et qui sont habituellement immergés à une profondeur variant entre 3 et 33 mètres. Généralement, les pêcheurs utilisent entre 20 et 100 filets qui forment de longs chapelets laissés sur place pendant deux ou trois jours avant d'être remontés à bord.
Les types d'engins utilisés et la période de la pêche à la lompe à Terre-Neuve constituent des facteurs clés qui influent sur le niveau de prises accessoires de phoques du Groenland. Les filets à grosses mailles utilisés pour la pêche constituent un obstacle important pour les phoques, qui ne peuvent peut-être pas les voir ou qui les attirent quand ils se remplissent de poissons. Dans certaines régions de la province, les activités de pêche empiètent sur les routes migratoires du phoque. Les jeunes phoques du Groenland nés à la fin de février et au début du mois de mars quittent les aires de mise bas se trouvant dans le sud du golfe du Saint-Laurent et la côte nord-est de Terre-Neuve en avril et en mai. Pendant cette migration printanière, certains phoques se déplacent le long des zones côtières de Terre-Neuve; ce sont eux qui se retrouvent pris dans les mailles des filets maillants installés pour les lompes.
Cette recherche fournit des estimations du nombre de phoques du Groenland prélevés annuellement entre 1970 et 2003. Les données proviennent d'un programme de surveillance des prises accessoires qui obligeaient les pêcheurs de lompes à consigner l'effort de pêche (les débarquements de rogues) et le nombre de phoques capturés quotidiennement entre 1989 et 2003. Avant 1989, les prises accessoires annuelles étaient estimées d'après les débarquements historiques de rogues.
Depuis le début de la pêche à la lompe en 1970 jusqu'en 1985, les prises accessoires de phoques du Groenland sont demeurées inférieures à 5 000 individus. En 1987, les prises se sont accrues, pour s'établir à un niveau record de 13 100 phoques, pour ensuite baisser à 3 600 individus en 1990. Les prises accessoires les plus élevées pour cette série chronologique s'élevaient à 46 394 phoques en 1994; les niveaux de capture sont demeurés supérieurs à 18 000 animaux jusqu'en 1997. En 2002 et 2003, on a observé un grand déclin de la pêche et les niveaux de prises accessoires ont baissé pour s'établir à moins de 10 000 animaux. Depuis 2003, les prises sont demeurées faibles. Ces estimations des prises accessoires annuelles varient selon plusieurs hypothèses clés, mais elles fournissent une vision à long terme des prises accessoires en tant que cause de mortalité.
4.3.2 Engin de pêche modifié pour réduire la mortalité accidentelle des cétacés
Edward Trippel

Câbles et engin de pêche modifiés
Photo: Edward Trippel
La mortalité accidentelle des cétacés attribuable aux prises accessoires et à l'enchevêtrement dans les engins de pêche commerciale est une source de préoccupation en matière de conservation dans le monde entier. On estime que plus de 10 % de la mortalité et plus de la moitié des blessures graves de la baleine noire de l'Atlantique Nord, une espèce extrêmement menacée, sont dues à son enchevêtrement dans les casiers à homards et les lignes des filets maillants. Depuis les années 1990, plusieurs techniques d'atténuation visant à réduire les mortalités de la population de baleines noires ont été mises à l'essai. L'utilisation de certaines de ces techniques dans la pêche commerciale est maintenant imposée par voie législative.
Toutefois, la population de baleines noires de l'Atlantique Nord demeure dans une situation précaire. Cette étude portait sur l'état actuel de développement et l'efficacité des techniques d'atténuation par l'ajout de sulfate de baryum au câble de l'engin de pêche.
La mise au point d'une ligne de fond à flottabilité neutre a été entreprise en vue de réduire les risques d'enchevêtrement de grosses baleines. Le produit qui en résulte est maintenu bien plus bas dans la colonne d'eau par rapport aux lignes de fond traditionnelles, mais dans les zones au fond dur, il était vulnérable au frottement et à l'usure. Pour réduire les mortalités une fois que les grandes baleines sont prises dans les filets, on a mis au point un câble de faible résistance recouvert de sulfate de baryum. La résistance à la rupture de ce produit était de 1 065 livres, ce qui est conforme aux limites imposées par la loi américaine, mais correspond à la moitié du poids des lignes de fond traditionnelles.
4.4 Relevés sismiques et sonars militaires
4.4.1 Regroupements de baleines boréales en quête de nourriture dans la mer de Beaufort canadienne et leur rôle dans l'atténuation des effets des bruits sous-marins sismiques
Lois Harwood

Baleines boréales vues d'un avion chargé des relevés
Photo: Lois Harwood
Un relevé aérien systématique par transects en travers du sud-est de la mer de Beaufort a été réalisé les 22 et 23 août 2007. L'objectif du relevé était de mettre à jour les connaissances sur la répartition des baleines boréales et leur utilisation de la mer de Beaufort, puisque le dernier relevé complet avait été effectué en 1986.
Vingt-quatre transects linéaires nord-sud ont été survolés, ce qui équivaut à environ 10 % de la couverture du relevé du sud-est de Beaufort. Les principaux observateurs ont observé 132 baleines boréales dans les transects, 38 à l'extérieur des transects et 71 pendant des vols de reconnaissance, pour un total de 241 baleines. Le nombre de baleines boréales observées dans les transects était plus de deux fois celui obtenu lors du relevé des années 1980, mais les zones générales où elles se rassemblaient étaient similaires.
Selon la définition de l'aire où se rassemblent des individus en quête de nourriture, soit plus de cinq baleines boréales par 100 kilomètres carrés, les baleines boréales étaient présentes dans trois grandes zones de regroupement d'individus en quête de nourriture en 2007, soit au large de la côte du Yukon entre Komakuk Beach et Shingle Point, près du rebord de la plateforme continentale au nord de l'estuaire du fleuve Mackenzie et au large de la péninsule de Tuktoyaktuk.
La tendance des baleines boréales à se regrouper et les connaissances en temps réel des zones où elles se regroupent permettent d'établir des procédures d'atténuation qui peuvent être
- plus restrictives dans les aires d'alimentation localisées des baleines boréales, pour éviter de perturber ou de blesser les baleines qui se nourrissent; et
- moins restrictives en dehors des aires d'alimentation, de façon à ne pas limiter de manière injustifiée l'activité de l'industrie. Des relevés aériens identiques sont prévus pour août 2008 et août 2009.
4.4.2 Surveillance acoustique des épaulards au large de la côte ouest du Canada
John Ford

Déploiement d'un dispositif d'enregistrement acoustique en mer
Photo: Hiromi Naito
On peut trouver l'épaulard – le plus grand prédateur marin – dans tous les océans, mais il est très rare dans la plupart des régions. Les eaux au large de la côte ouest du Canada abritent une abondance exceptionnelle de cette espèce, qui devient une espèce « vedette » de l'environnement marin sauvage de la Colombie-Britannique. Depuis longtemps, les épaulards font l'objet de recherches sur le terrain par des scientifiques spécialistes des mammifères marins de la Station biologique du Pacifique (SBP) du MPO. Chaque année depuis le début des années 1970, on étudie les épaulards au moyen de la photo-identification des individus à partir des marques naturelles. La Station tient des registres de plus de 700 baleines provenant de trois assemblages distincts – ou écotypes – d'épaulards dans les eaux de la Colombie-Britannique. Cette surveillance annuelle de la population est essentielle aux efforts de rétablissement pour ces écotypes, qui sont tous répertoriés en vertu de la Loi sur les espèces en péril du Canada.
Grâce à cette étude à long terme, de nombreux aspects du cycle biologique et de l'écologie des épaulards en Colombie-Britannique nous sont maintenant connus, mais nos connaissances comportent encore de nombreuses lacunes. Une grande source d'incertitude a trait aux lieux de séjour des épaulards en hiver, quand la plupart des baleines quittent leurs habitats estivaux dans les eaux côtières protégées autour de l'île de Vancouver. Trouver des épaulards dans les eaux éloignées au large de la côte ouest en hiver constitue un défi de taille, et les petites embarcations que nous utilisons pour nos études en été ne sont pas à la hauteur de cette ambition. Le marquage des baleines au moyen d'étiquettes émettrices n'est pas encore une option viable en raison de la difficulté de fixer l'étiquette aux baleines sans les capturer et du manque de solidité de ces étiquettes, qui ne durent qu'un mois ou deux.
Pour en savoir plus sur la répartition hivernale des épaulards, nous établissons un réseau de stations de surveillance acoustique sous-marine éloignées à des endroits stratégiques le long de la côte de la Colombie-Britannique pour localiser les baleines grâce à leurs chants. Les épaulards conviennent parfaitement à ce genre de surveillance, puisque leurs chants sont fortement stéréotypés et diffèrent entre les populations et les groupes sociaux. Ainsi, avec seulement quelques minutes de chants d'un épaulard, nous pouvons déterminer son écotype et, dans le cas des épaulards « résidents », nous pouvons identifier les clans et les groupes concernés.
Nos stations de surveillance acoustique sont de deux types différents: côtières et extracôtières. Un prototype de station côtière a été mis à l'essai pendant quatre hivers à l'île Langara, au large de l'extrémité nord-ouest des îles de la Reine-Charlotte, ou Haida Gwaii. Cette station comprend une boîte étanche contenant un ordinateur spécialisé doté d'un système de détection des sons établi sur mesure. Il est alimenté par des batteries d'accumulateurs qui demeurent chargées au moyen de panneaux solaires. L'instrument est relié par un câble armé à un microphone sous-marin, ou hydrophone, placé sur le plancher océanique à environ 100 mètres de la côte. Quand l'hydrophone capte des sons de baleine, le système de détection acoustique enclenche l'ordinateur, qui enregistre un échantillon de sons sous-marins sur un disque dur à des fins d'analyse ultérieure. Cette station a livré de nombreuses données nouvelles sur la présence des baleines à cet endroit éloigné, et nous espérons déployer d'autres stations côtières à l'avenir.

Station d'enregistrement acoustique à distance (Orcabox) à l'île Langara
Photo: John Ford
Notre deuxième type de station de surveillance a une fonction similaire aux unités côtières, mais l'ensemble du système est placé dans une boîte étanche submersible qui doit être déployée au large. La boîte contient un hydrophone, un système informatique miniaturisé avec un disque dur pour stocker les enregistrements numériques et des batteries qui permettent une surveillance et des enregistrements pendant un maximum d'un an. Les stations extracôtières sont ancrées au fond marin et suspendues à 50 mètres sous la surface par des flotteurs. Elles sont récupérées au moyen d'une libération acoustique, qui déconnecte l'ancre et permet à la station de flotter à la surface en vue de sa récupération.
La troisième méthode de surveillance acoustique des épaulards est devenue possible grâce à notre collaboration avec deux initiatives menées au large de la côte ouest – VENUS (Réseau expérimental sous-marin de Victoria) et NEPTUNE (Northeast Pacific Time-Series Underwater Networked Experiments). VENUS et NEPTUNE comportent tous deux des nœuds d'échantillonnage océanographique sur le plancher océanique qui sont reliés à Internet par un câble à fibres optiques. Ces nœuds comprennent des hydrophones optimisés pour la surveillance des sons des baleines et ils permettront aux chercheurs et au public d'écouter des épaulards en temps réel.
À mesure que notre réseau de surveillance acoustique des épaulards prend de l'expansion, il devrait continuer de fournir de nouvelles données sur les habitudes migratoires saisonnières de ces prédateurs énigmatiques.
4.5 Les mammifères marins comme sentinelles de la contamination environnementale
4.5.1 Évaluation des effets éventuels de la prospection de gisements d'hydrocarbures près des côtes sur les phoques annelés dans la région de la mer de Beaufort
Lois Harwood

Phoque annelé à un trou d'air
Photo: DFO
Les objectifs de cette étude étaient de cerner et d'évaluer les répercussions possibles des activités industrielles océaniques sur les populations de phoques résidentes, en vue de formuler des avis sur les mesures d'atténuation et les études de surveillance qui pourraient être employées efficacement à l'avenir. L'étude portait sur le phoque annelé entre 2003 et 2006 dans l'habitat de banquise côtière au nord de l'estuaire du fleuve Mackenzie dans la mer de Beaufort.
Le site de forage Paktoa 2006 de Devon Canada se trouvait près du bord de la banquise, qui constitue une caractéristique très importante de l'écosystème marin arctique et est essentiel pour les phoques. Elle est utilisée pour établir et défendre les territoires d'accouplement, donner naissance aux petits et en prendre soin, se reproduire et se reposer. Pendant les trois premières années de l'étude, l'activité industrielle n'avait pas encore commencé à Paktoa, tandis que la quatrième année, l'étude a eu lieu pendant la dernière partie de la saison de forage exploratoire. En 2006, Devon a terminé le forage et a abandonné le puits.
La distance moyenne entre les tanières et les trous d'aération, d'une part, et Paktoa, d'autre part, n'a guère changé au cours de la période et il en va de même pour les structures gelées utilisées activement par les phoques, où l'on n'a observé aucune différence significative entre l'année d'activité industrielle et les années précédentes. En 2006, la distribution statistique des distances des structures utilisées par les phoques par rapport à la route de glace, à la piste d'atterrissage et au campement de recherche n'a montré aucune tendance à l'évitement ou à l'attrait de ces sites.
Vingt phoques annelés ont été capturés, munis d'émetteurs satellites et relâchés près du site de Paktoa. On n'a pas observé de différences statistiques notables dans les déplacements et la taille des territoires fréquentés par ces animaux marqués entre 2005 et 2006, quand l'état des glaces était fort différent. Par ailleurs, 68 phoques annelés ont été prélevés afin d'examiner leur état corporel et l'état reproducteur de la population locale. Les spécimens prélevés avaient un bon état corporel et d'importantes réserves de gras; leur état reproducteur était normal et la plupart avaient des proies dans l'estomac.
Dans l'ensemble, l'étude a livré d'importantes données de référence sur l'utilisation par les phoques annelés des eaux à proximité des côtes de la mer de Beaufort au printemps et sert de référence à toute étude ultérieure concernant plusieurs activités de forage ou encore des activités de longue durée. Les résultats donnent à penser qu'une saison de forage industriel dans le site de Paktoa n'a pas eu d'effet perceptible sur les phoques annelés dans l'aire à l'étude. Les effets d'expositions plus longues à cette activité industrielle demeurent inconnus.
4.5.2 Les épaulards comme indicateurs de la pollution en milieu océanique
Peter Ross

Épaulards devant une raffinerie de pétrole
Photo: Ingrid Visser
Malgré des efforts internationaux visant à lutter contre les biphényles polychlorés (BPC) et de nombreux autres contaminants, ceux-ci demeurent présents dans l'environnement et peuvent présenter des risques pour la santé des mammifères marins se trouvant en haut de la chaîne trophique. Les populations d'épaulards de la Colombie-Britannique font face à de nombreuses pressions. Le Comité sur le statut des espèces menacées de disparition au Canada classe les résidents du sud comme étant en danger de disparition et les résidents du nord et les épaulards de passage comme étant menacés. Les épaulards résidents consomment presque exclusivement du poisson, tandis que les populations de passage consomment presque exclusivement des mammifères marins.
Plusieurs facteurs nuisent au rétablissement des épaulards résidents de la Colombie-Britannique, dont la petite taille de la population, la raréfaction de leur proie principale (saumon), le bruit et la perturbation. En outre, les épaulards résidents sont une cible d'observation mythique pour les touristes, et des mesures sont prises pour assurer que les activités d'écotourisme n'entravent pas les processus vitaux normaux, entre autres l'alimentation, la socialisation et la reproduction.
Le phoque commun a permis de nous éclairer sur la menace que représentent les contaminants comme les BPC pour les épaulards. Ce petit mammifère marin non migrateur a une physiologie similaire à celle de l'épaulard. Son échantillonnage entraîne moins de perturbations que la réalisation d'autres biopsies sur les épaulards à l'aide de dards. L'abondance des phoques communs, qui sont au nombre de 130 000 en Colombie-Britannique, permet aux chercheurs de choisir les animaux d'un même âge et d'un état semblable, ce qui fournit des données plus fiables.
Que la source des contaminants soit située à proximité ou très éloignée, les épaulards de la Colombie-Britannique en accumulent dans leur organisme en consommant des proies côtières. Le saumon, principale source de nourriture des épaulards, migre des océans aux milieux côtiers, transportant du même coup de petites quantités de produits toxiques. Les espèces comme le sébaste, qui passent leur vie plus près des côtes, ont également une teneur plus élevée en contaminants.
La position des proies dans la chaîne trophique est également importante. Les épaulards nomades se nourrissent de phoques, d'otaries, de marsouins et d'autres mammifères qui ont déjà accumulé des toxines en ingérant des organismes des niveaux inférieurs de la chaîne trophique. Ainsi, les épaulards nomades risquent plus d'être touchés par les effets toxiques que leurs cousins résidents qui se nourrissent de poissons. Les épaulards de la Colombie-Britannique aident les chercheurs et les gestionnaires à mieux comprendre le sort des contaminants environnementaux à l'échelle mondiale.
4.6 Événements d'échouement et de mortalité
4.6.1 Mortalité causée par la chasse chez les phoques du Groenland dans l'Atlantique Nord-Ouest
Garry Stenson

Navires phoquiers qui partent chasser le phoque du Groenland près des îles de la Madeleine
Photo: Mike Hammill
On a besoin d'information sur le niveau de mortalité causée par l'homme pour établir des estimations précises de toute population. Ceci est particulièrement important pour les espèces exploitées qui font l'objet de prises importantes, et cette information est donc nécessaire pour élaborer un plan de gestion responsable. La population de phoques du Groenland de l'Atlantique Nord-Ouest est sujette à divers types de mortalité causée par l'homme, ente autres la pêche de subsistance au Groenland et dans l'Arctique canadien, les prises commerciales dans les eaux méridionales du Canada, les animaux tués mais non débarqués et les prises accessoires dans les engins de pêche commerciale.
L'objectif de cette étude était de résumer les estimations disponibles et de les mettre à jour pour la période 1952-2004. Les pêches de subsistance et commerciale représentaient la majorité des prélèvements. Entre 1952 et 1971, les prises se sont établies en moyenne à plus de 288 000 phoques pour la chasse commerciale. Les quotas ont été mis en place en 1972, et jusqu'à la fin de la chasse à bord de gros bateaux en 1982, 165 000 phoques étaient prélevés chaque année. Après 1982, les prises se sont élevées en moyenne à 52 000, jusqu'en 1995, où elles ont augmenté pour atteindre une moyenne de 258 000 phoques.
Au Groenland, les prises étaient inférieures à 20 000 avant 1980, puis ont augmenté progressivement pour atteindre un sommet de plus de 100 000 en 2002 et ont diminué récemment pour s'établir à quelque 70 000 phoques. On dispose de données limitées sur les prises dans l'Arctique canadien, mais il semble que les nombres soient relativement faibles (moins de 5 000). Ainsi, le total des prélèvements entre 1952 et 1982 s'établissait à 388 000, mais il a chuté à 178 000 annuellement entre 1983 et 1995. Le Canada et le Groenland ont augmenté leurs prises à partir de 1996, ce qui a donné lieu à un prélèvement annuel moyen de 471 000 individus. Les jeunes de l'année représentent environ 68 % des prélèvements actuels.
4.6.2 Bélugas de l'estuaire du Saint-Laurent – Examen du programme des carcasses
Lena Measures

Examen d'un béluga échoué
Photo: Lena Measures
En 2005, un atelier sur le programme des carcasses de bélugas de l'estuaire du Saint-Laurent s'est tenu à l'Institut Maurice-Lamontagne. Quelque 25 scientifiques et gestionnaires participant au programme ou intéressés par le programme et des consultants invités y ont pris part. L'atelier avait une double finalité: 1) évaluer l'utilité du programme en fonction de quatre thèmes principaux, soit les causes de mortalité et les maladies, les contaminants et les effets toxiques, la biologie et la démographie, et la conservation; et 2) déterminer l'avenir du programme en examinant les options et les conséquences. On a présenté l'historique du programme de récupération des carcasses ainsi qu'une description du programme de nécropsie. D'autres exposés ont résumé les connaissances actuelles sur les maladies parasitaires et infectieuses, l'exposition aux contaminants et leurs effets toxiques, l'information écologique et démographique et les mesures de conservation (parc marin national et aire marine protégée proposée) conçues pour protéger le béluga et son habitat. Un dernier exposé sur l'utilité d'un programme d'intervention auprès des mammifères marins échoués en Californie a été présenté à des fins de comparaison.
Quatre recommandations clés ont été formulées à l'issue de l'atelier :
- On doit maintenir le programme de carcasses de bélugas de l'estuaire du Saint-Laurent, une étude de cas reconnue à l'échelle mondiale;
- il faudrait communiquer et intégrer les données disponibles, normaliser et documenter les protocoles et coordonner les banques de tissus et les bases de données archivées, etc.
- les volets de base du programme (surveillance, nécropsie et échantillonnage) doivent se poursuivre et bénéficier d'un financement adéquat; et
- des projets de recherche spéciaux devraient être financés pour répondre à des questions précises soulevées par les résultats du programme de base de même qu'à des besoins de gestion particuliers. On a jugé essentiel de valider, d'examiner et d'intégrer plus de 20 années de données consultables dans une base de données accessible de sorte à pouvoir vérifier diverses hypothèses et à les utiliser pour mieux comprendre pourquoi cette population ne s'est pas rétablie. On peut consulter un compte rendu (2007-005) de l'atelier à l'adresse http://www.meds-sdmm.dfo-mpo.gc.ca/csas/applications/Publications/publicationIndex_f.asp.
4.6.3 Réduction du risque de collision entre les navires et les baleines noires dans la région du bassin Roseway du sud-ouest de la plateforme Scotian
Heath Stone

Baleine noire femelle et son baleineau
Photo: NOAA-NMML
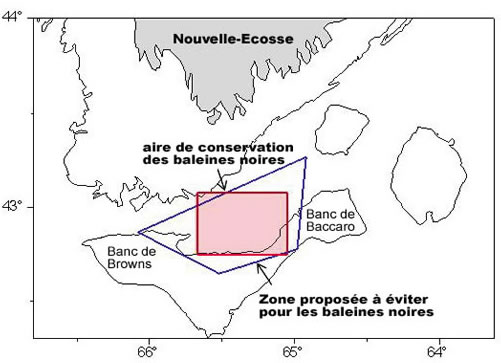
Figure 10. Le sud-est de la Plateforme Scotian avec l'emplacement de l'aire de conservation de la baleine noire du bassin Roseway (rouge) et la zone désignée à éviter par la navigation commerciale (bleu).
La baleine noire de l'Atlantique Nord est l'une des plus grandes baleines. Elle est également l'une les plus en danger et est répertoriée comme espèce menacée de disparition dans l'annexe 1 de la Loi sur les espèces en péril du Canada. On estime actuellement la population de baleines noires à 350-400 individus. L'abondance de l'espèce semblait diminuer pendant les années 1990, et la mortalité causée par l'homme qui a été documentée est principalement attribuable aux collisions avec des bateaux. En 1993, le MPO a désigné deux aires de conservation: le bassin de Grand Manan (baie de Fundy) et le bassin Roseway (entre le banc de Browns et le banc de Baccaro). Les deux aires sont importantes pour l'alimentation et la socialisation des baleines et les concentrations de baleines noires y sont élevées en été et à l'automne.
Des mesures de conservation visant à réduire les risques de collision entre baleines noires et navires dans l'aire de conservation du bassin de Grand Manan ont été mises en œuvre avec succès en 2003 par Transports Canada et le MPO. En dehors de la baie de Fundy, le bassin Roseway est la seule autre aire connue en eaux canadiennes où les baleines noires de l'Atlantique Nord se réunissent en nombre relativement élevé, mais il n'existe actuellement aucune mesure reconnue de gestion du trafic maritime dans cette région.
En 2006, le Comité national d'examen par les pairs sur les mammifères marins du Secteur des sciences du MPO a passé en revue une proposition visant à désigner une « zone à éviter » autour de l'aire de conservation du bassin Roseway (figure 10). La proposition reposait sur des données scientifiques, qui comparaient la répartition des baleines noires avec l'information sur le trafic maritime traversant la région. Le Comité a conclu qu'une réduction du trafic maritime dans cette région donnerait lieu à une réduction des risques de collision entre les baleines noires et les navires et accroîtrait donc les chances de rétablissement de l'espèce.
Le 7 octobre 2007, l'Organisation maritime internationale (OMI) a adopté une proposition (présentée par Transports Canada) en vue de soustraire à la navigation 1 780 kilomètres carrés du bassin Roseway, en les désignant « zones à éviter » par les navires de 300 tonnes de jauge ou plus, seulement en transit, entre le 1er juin et le 31 décembre. La date d'entrée en vigueur est le 1er mai 2008.
4.7 Changement climatique
4.7.1 Les phoques annelés comme indicateurs du changement
Lois Harwood

Remise en liberté d'un phoque annelé marqué dans son trou
Photo: Lois Harwood
La répartition du phoque annelé est circumpolaire, et ce phoque est l'espèce de pinnipède la plus abondante dans l'océan Arctique. Il est considéré comme un indicateur raisonnable de la santé de l'écosystème en raison de sa position en haut de la chaîne trophique. Les phoques annelés sont omniprésents et constituent d'importantes proies pour les ours polaires; les changements dans la répartition et l'abondance du phoque peuvent avoir des conséquences à long terme sur la survie des ours polaires. Les phoques annelés constituent une ressource de subsistance précieuse pour les Inuvialuits. Les recherches menées auparavant sur les phoques annelés de l'ouest de l'Arctique ont révélé une tendance à la baisse dans l'abondance de l'espèce, une réduction des taux d'ovulation chez les femelles adultes, une réduction du nombre de nouveau-nés dans la pêche de subsistance et une réduction du nombre de sites propices de mise bas en association avec les conditions de glace épaisse de l'hiver 1974-1975.
Des échantillons de phoques ont été prélevés pendant la chasse de subsistance par un chasseur de phoques hautement expérimenté d'Ulukhaktok, dans les Territoires du Nord-Ouest. Les paramètres examinés incluent la détermination du taux d'ovulation des femelles adultes, la proportion de nouveau-nés dans la chasse de subsistance, l'importance des classes d'âge, l'état corporel (épaisseur du petit lard, indice de masse corporelle) des phoques capturés et l'enregistrement d'observations et d'événements inhabituels par les pêcheurs. Les taux d'ovulation sont toujours demeurés élevés entre 1992 et 1999 et ont baissé légèrement récemment. La proportion annuelle de nouveau-nés dans la pêche en mer ouverte a également affiché une tendance à la baisse entre 2000 et 2006. L'épaisseur du petit lard a diminué entre 1992 et 1996 pour tous les groupes d'âge, à l'exception des jeunes qui viennent d'être sevrés.
Cette tendance à la baisse dans la reproduction des phoques et leur état corporel suivait de près le nombre de jours d'eau libre en 2000 et en 2002-2005. Lorsque les phoques annelés passent plus de temps en eau libre, comme ce fut le cas en 1998 et en 2001, ils sont plus gras et la production de nouveau-nés est élevée. On ne comprend pas bien les mécanismes, mais ils semblent liés aux changements dans le moment de la dislocation de la glace de mer au printemps. Le moment du dégel influe sur le moment de l'établissement de conditions océanographiques qui favorisent la production d'aliments pour les phoques, comme la morue.
4.7.2 Épaulards et changement climatique
Steve Ferguson

Sauts d'observation des épaulards
Photo: Graeme Ellis
Bien que ceci soit l'objet d'une controverse, les épaulards ont joué un rôle en tant que modificateurs importants de l'écosystème créant des cascades trophiques par suite de perturbations de l'écosystème, notamment la chasse commerciale à la baleine. Les modèles de changements climatiques prédisent des réductions importantes de la banquise, ce qui entraînera probablement des changements dans la répartition et l'abondance des espèces. On a émis l'hypothèse que les épaulards migreront dans les eaux de l'Arctique à mesure que la banquise disparaîtra, et qu'en tant que prédateurs d'un niveau trophique supérieur, ils déclencheront d'importants ajustements écosystémiques. Dès lors, on prévoit que les épaulards joueront un rôle de premier plan dans la réorganisation de l'océan Arctique qui subit actuellement les effets du réchauffement climatique.
Dans cette étude, les changements dans la répartition des épaulards dans la baie d'Hudson ont été mesurés avec la diminution de la banquise comme example servant à illustrer les réajustements globaux qui se produisent par suite de l'évolution du climat. On a résumé les données sur les épaulards dans la baie d'Hudson, le détroit d'Hudson et le bassin Foxe et établi un lien avec une série de données historiques sur la banquise. Les résultats révèlent l'existence de « goulots d'étranglement », où la banquise entrave les déplacements des épaulards et restreint leur répartition dans l'Arctique. Le détroit d'Hudson semble avoir été un important goulot d'étranglement de glace marine qui s'est ouvert il y a 50 ans, permettant l'apparition des épaulards et la progression de la répartition d'un prédateur évitant la glace.
Les observations d'épaulards ont augmenté de manière exponentielle et on en signale maintenant dans la région de la baie d'Hudson chaque été. La disparition d'autres goulots d'étranglement est à prévoir en raison de la fonte continue de la banquise, ce qui entraîne des cascades trophiques prédateur-proie intermittentes dans l'Arctique.
5.0 Publications de 2006-2008
- Anderson, J.T., D.V. Holliday, R. Kloser, D. Reid et Y. Simard. « Acoustic seabed classification of marine physical and biological landscapes », dans Anderson et coll. (sous la dir. de). Rapport des recherches collectives du CIEM, nº 286, 2007, 183 pp.
- Anderson, J.T., D.V. Holliday, R. Kloser, D. Reid et Y. Simard. « Future directions for acoustic seabed classification science », dans Anderson et coll. (sous la dir. de). « Acoustic seabed classification of marine physical and biological landscapes », dans Rapport des recherches collectives du CIEM, nº 286, 2007, 183 pp.
- Anderson, J.T., D.V. Holliday, R. Kloser, D. Reid et Y Simard. « Acoustic seabed classification: Current practice and future directions », dans ICES Journal of Marine Science, nº 65, 2008, doi:10.1093/icesjms/fsn061.
- Assuncao M.G.L., K.A. Miller, N.J. Dangerfield, S.M. Bandiera et P.S. Ross. « Cytochrome P450 1A expression and organochlorine contaminants in harbour seals: Evaluating a biopsy approach », dans Comparative Biochemistry and Physiology C-Toxicology and Pharmacology, vol. 145, nº 2, 2007, p. 256-264.
- Austin, D., W.D. Bowen, J.I. McMillan et D.J. Boness. « Stomach temperature telemetry reveals temporal patterns of foraging success in a free-ranging marine mammal », dans Journal of Animal Ecology, vol. 75, 2007, p. 408-420.
- Austin, D., W.D. Bowen, J.I. McMillan et S.J Iverson. « Linking movement, diving and habitat to feeding a large marine predator », dans Ecology, vol. 87, 2007, p. 3095-3108.
- Bahoura, M., R. Lepage, X. Mouy, C.W. Park, J. Rouat, N. Ray, D. Seebaruth, Y. Simard et M. Sirois. « Development and experimentation of a satellite buoy network for real-time acoustic localization of whales in the St. Lawrence », compte rendu d'OCEANS'06 MTS/IEEE – Boston, IEEE, nº de cat. 06CH37757C, ISBN: 1-4244-0115-1. IEEE, Piscataway, NJ, USA, 2006.
- Bahoura, M. et Y. Simard. « Blue whale calls characterization using chirplet transform », dans Journal of the Acoustical Society of America, vol. 123, 2008, p. 3779.
- Bahoura, M. et Y. Simard. « Chirplet transform applied to simulated and real blue whale (Balaenoptera musculus) calls », dans Image and Signal Processing in Lecture Notes in Computer Science, nº 5099, p. 296-303. https://link.springer.com/content/pdf/10.1007%2F978-3-540-69905-7_34.pdf, 2008.
- Beck, C.A., S.J. Iverson, W.D. Bowen et W. Blanchard. « Sex differences in grey seal diet reflect seasonal variation in foraging behaviour and reproductive expenditure: evidence from quantitative fatty acid signature analysis », dans Journal of Animal Ecology, nº 76, 2007, p. 490-502.
- Bedard, C. et Y. Simard. « Automatic detection of white whale vocalizations in St. Lawrence estuary and occurrence pattern. Proceedings of the Canadian Acoustical Society Annual Meeting », dans Acoustique canadienne, vol. 34, nº 3, 2006, p. 84-85.
- Benjamins, S., J.Lawson et G. Stenson. « Recent harbour porpoise bycatch in gillnet fisheries in Newfoundland and Labrador », dans Journal of Cetacean Research and Management, vol. 9, 2007, p. 189-199.
- Benoit, D., Y. Simard et L. Fortier. « Hydro-acoustic detection of large winter aggregations of Arctic cod (Boreogadus saida) at depth in ice-covered Franklin Bay (Beaufort Sea) », dans Journal of Geophysical Research, vol. 113, 2008, C06S90, doi:10.1029/2007JC004276.
- Boily, F., S. Beaudoin et L.N. Measures. « Hematology and serum chemistry of harp (Phoca groenlandica) and hooded seals (Cystophora cristata) during the breeding season, in the Gulf of St. Lawrence, Canada », dans Journal of Wildlife Diseases, vol. 2, 2006, p. 115-132.
- Boness, D.J., W.D. Bowen, B. Bulheier et G. Marshall. « Mating tactics and mating system of an aquatic mating pinniped: the harbour seal », dans Behavioural Ecology and Sociobiology, vol. 61, 2006, p. 119-130.
- Bowen, W.D., C.A. Beck, S.J. Iverson, D. Austin et J.I. McMillan. « Linking predator foraging behaviour and diet with variability in continental shelf ecosystems: grey seals of eastern Canada », dans Boyd, I., Wanless, S. et Camphuysen, C.J. (sous la dir. de). Top Predators in marine ecosystems: their role in monitoring and management, Cambridge University Press, Cambridge, UK, 2006, p. 63-81.
- Bowen, W.D., S.J. Iverson, J.I. McMillan et D.J. Boness. « Reproductive performance in grey seals: age-related improvement and senescence in a capital breeder », dans Journal of Animal Ecology, vol. 75, 2006, p. 2319-2326.
- Bowen, W.D. et G. Harrison. « Seasonal and interannual variability in grey seal diets on Sable Island, eastern Scotian Shelf », dans T. Haug, M. Hammill et D. Olafsdottir (sous la dir. de). Grey seals in the North Atlantic and the Baltic, Nammco scientific publications, vol 6, 2007, 227 p. The North Atlantic Marine Mammal Commission. Tromso, Grafisk Nord, Finnsnes, p. 123-134.
- Bowen, W. D., D.J. Boness et L.L. Lowry. Review of the 2007 Draft Steller Sea Lion Recovery Plan for the North Pacific Fishery Management Council, juillet 2007, 25 p.
- Bowen, W.D., J.I. McMillan et W. Blanchard. « Reduced rate of population growth of grey seals at Sable Island: evidence from pup production and age of primiparity », dans Marine Mammal Science, vol. 23, 2007, p. 48-64.
- Bowen, W.D, J.I. McMillan, D. Lidgard et W. Blanchard. Diminution continue du taux de croissance de la population de phoques gris à l'île de Sable, document de recherche 2007/087 du Secrétariat canadien de consultation scientifique, 2007
- Bowen, W. D., L. Gage, D. Goodman et L.L. Lowry. Report of the Independent Review Panel on the National Marine Fisheries Service's Implementation of the Permit Program for Research on Steller Sea Lions and Northern Fur Seals, 2008, 48 p.
- Bowen, W. D., M.O. Hammill, M. Koen-Alonso, G. Stenson, D.P. Swain et K. Trzcinski. Compte rendu de l'atelier national sur les répercussions des phoques sur les populations de poissons dans l'est du Canada (1re partie), compte rendu 2008/021 du Secrétariat canadien de consultation scientifique, 2008.
- Breed, G.A., W.D. Bowen, J.I. McMillan et M.L. Leonard. Sexual segregation of seasonal foraging habitats in a non-migratory marine mammal, compte rendu de la Royal Society B-Biological Sciences, vol. 273, nº 1599, 2006, p. 2319-2326.
- Buren, A.D., M. Koen-Alonso et G.B. Stenson. « Predator-prey interaction between harp seals and Atlantic cod: An exploration of sources of variation », dans Report of the Workshop on the Decline and Recovery of cod Stocks throughout the North Atlantic, including tropho-dynamic effects (WKDRCS), ICES CM 2006/OCC, vo. 12, 2006, p. 151-152.
- Burns, J.M., K.C. Lestyk, L.P. Folkow, M.O. Hammill et A.S. Blix. « Size and distribution of oxygen stores in harp and hooded seals from birth to maturity », dans Journal of Comparative Physiology B – Biochemical Systemic and Environmental Physiology, vol. 177, nº 6, 2007, p. 687-700.
- Calambokidis, J., E.A. Falcone, T.J. Quinn, A.M. Burdin, P.J. Clapham, J.K.B. Ford, C.M. Gabriele, R. LeDuc, D. Mattila, L. Rojas-Bracho, J.M. Straley, B.L. Taylor, R.J. Urbán, D. Weller, B.H. Witteveen, M. Yamaguchi, A. Bendlin, D. Camacho, K. Flynn, A. Havron, J. Huggins et N. Maloney. SPLASH: Structure of Populations, Levels of Abundance and Status of Humpback Whales in the North Pacific, rapport final pour le contrat AB133F-03-RP-00078, Cascadia Research, Olympia, WA, 2008.
- Calambokidis, J., J. Barlow, J., J.K.B. Ford, T.E. Chandler et A.B. Douglas. Sous presse. « Insights into the population structure of blue whales in the eastern North Pacific from recent sightings and photographic identification », dans Marine Mammal Science.
- Chambellant, M. et S.H. Ferguson. Sous presse. « Ageing live ringed seals (Phoca hispida): which tooth to pull? », dans Marine Mammal Science.
- Chassot, E., A. Caskenette, D. Duplisea, M. Hammill, H. Bourdages, Y. Lambert et G. Stenson. Un modèle de prédation des phoques du Groenland (Phoca groenlandica) sur le stock de morues (Gadus morhua) du nord du golfe du Saint-Laurent, document de recherche 2007/066 du Secrétariat canadien de consultation scientifique, 2007.
- Chassot, E., D. Duplisea, M. Hammill, A. Caskenette, N. Bousquet, Y. Lambert et G. Stenson. Sous presse. « The role of predation by harp seals (Pagophilus groenlandicus) in the collapse and non-recovery of northern Gulf of St. Lawrence cod (Gadus morhua) », dans Marine Ecology Progress Series.
- Chikoski, J., S.H. Ferguson et L. Meyer. « Effects of water addition on soil arthropods and soil characteristics in a precipitation-limited environment », dans Acta Oecologia, vol. 30, 2006, p. 203-211.
- Clark, C.A., J.M. Burns, J.F. Schreer et M.O. Hammill. « Erythropoietin concentration in developing harbour seals », dans General and Comparative Endocrinology, vol. 147, nº 3, 2006, p. 262-267.
- Clark, CA, J.M. Burns, J.F. Schreer et M.O. Hammill. « A Longitudinal And Cross-Sectional Analysis Of Total Body Oxygen Store Development », dans « Nursing Harbor Seals (Phoca vitulina) », Journal of Comparative Physiology B, vol. 177, 2007, p. 217-227.
- Coltman, D.W., G. Stenson, M.O. Hammill, T. Haug, S. Davis et T.L. Fulton. « Panmictic population structure in the hooded seal (Cystophora cristata) », dans Molecular Ecology, vol. 16, nº 8, 2007, p. 1639-1648.
- Cote, D., H, Stewart, R.S. Gregory, J. Gosse, J. Reynolds, G.B. Stenson et E.H. Miller. « Prey selection by marine –coastal river otters (Lontra canadensis) in Newfoundland, Canada », dans Journal of Mammalogy, vol. 89, 2008, p. 1001-1011.
- Cosens, S.E., H. Cleator et P. Richard. Nombre de baleines boréales (Balaena mysticetus) dans l'est de l'Arctique canadien, d'après les relevés aériens d'août 2002, 2003 et 2004, document de recherche 2006/052 du Secrétariat canadien de consultation scientifique, 2006.
- Cyr, C., J.-F. Gosselin, M.O. Hammill, V. Lesage, N. Roy et Y. Simard. Impacts de la construction d'un port méthanier à Gros-Cacouna sur les mammifères marins, réponse des Sciences 2007-010 du Secrétariat canadien de consultation scientifique du MPO, 2007.
- Davidson, F.J.M., M.O. Hammill et G. Stenson. « Expanding the gathering of real-time in situ oceanographic data with the DFO Marine Mammal Research Program », dans le Bulletin du PMZA, vol. 6, 2007, p. 63-68.
- Dietz, R., A.D. Shapiro, M. Bakhtiari, J. Orr, P.L. Tyack, P. Richard, I. Grønborg Eskesen et G. Marshall. « Upside-down swimming behaviour of free-ranging narwhals », dans BMC Ecology, vol. 7, nº 14, 2007, doi:10.1186/1472-6785-7-14.
- Dietz, R., M.P. Heide-Jørgensen, P. Richard, J. Orr, K. Laidre et H.C. Schmidt. « Movements of narwhals (Monodon monoceros) from Admiralty Inlet monitored by satellite telemetry », dans Polar Biology, 2008, DOI 10.1007/s00300-008-0466-4.
- Dueck, L.P., M.P. Heide-Jorgensen, M.V. Jensen et L.D. Postma. Mise à jour des études sur les déplacements des baleines boréales (Balaena mysticetus) dans l'est de l'Arctique, de 2003 à 2005, grâce à la télémétrie par satellite, document de recherche 2006/050 du Secrétariat canadien de consultation scientifique du MPO, 2006, 25 p.
- Dueck, L. et P. Richard. Preliminary evaluation of selective hunting and recovery of Eastern Canadian Arctic-West Greenland bowhead whales. document de recherche 2008/xx du Secrétariat canadien de consultation scientifique, 2008.
- Dueck, L., P. Richard et S. Cosens. Examen et nouvelle analyse de l'évaluation par Cosens et coll. (2006) des relevés aériens de l'abondance du stock de baleines boréales de l'est de l'Arctique canadien, document de recherche 2007/080 du Secrétariat canadien de recherche consultative, 2008.
- Ferguson, S.H. « The Influences of environment, mating habitat, and predation on evolution of pinniped lactation strategies », dans Journal of Mammalian Evolution, vol. 13, 2006, p. 63-82.
- Ferguson, S.H. et J.W. Higdon. « How seals divide up the world: environment, life-history, and conservation », dans Oecologia, vol. 150, 2006, p. 318-329.
- Ferguson, S.H., J. W. Higdon et S. Larivière. « Does seasonality explain the evolution and maintenance of delayed implantation in the family Mustelidae (Mammalia: Carnivora)? », dans Oikos, vol. 114, 2006, p. 249-256.
- Ferguson, S.H. et S. Larivière. « How social behaviour links environment and body size in mammalian carnivores », dans Open Journal of Ecology, vol. 1, 2008, p. 1-7.
- Ford, J.K.B. Évaluation des habitats essentiels des populations résidentes d'orques dans les eaux de la côte du Pacifique du Canada, document de recherche 2006/072, Secrétariat canadien de consultation scientifique, ministère des Pêches et des Océans, Ottawa, Ontario, 2006.
- Ford, J.K.B. et G.M. Ellis. « Selective foraging by fish-eating killer whales in British Columbia », dans Marine Ecology Progress Series, vol. 316, 2006, p. 185-199.
- Ford, J.K.B., G.M. Ellis et J.W. Durban. Évaluation du potentiel de rétablissement de l'épaulard migrateur du Pacifique Nord-Est vivant dans les eaux côtières de la Colombie-Britannique, document de recherche 2007/088 du Secrétariat canadien de consultation scientifique, ministère des Pêches et des Océans, Ottawa, Ontario, 2007.
- Ford, J.K.B. et R.R. Reeves. « Fight or flight: antipredator strategies of baleen whales », dans Mammal Review, vol. 38, 2008, p. 50-86.
- Ford, J.K.B. Sous presse. « Killer whale Orcinus orca », dans W.F. Perrin, B. Wursig et H.G.M. Thewissen (sous la dir. de). The Encyclopedia of Marine Mammals Second Edition, Elsevier, New York, NY.
- Ford, J.K.B. Sous presse. « Dialects », dans W.F. Perrin, B. Wursig et H.G.M Thewissen (sous la dir. de). The Encyclopedia of Marine Mammals Second Edition, Elsevier, New York, NY.
- Ford, J.K.B. et G.M. Ellis. Sous presse. « You are what you eat: ecological specializations and their influence on the social organization and behaviour of killer whales », dans J. Yamagiwa et L. Karczmarski (sous la dir. de). Primates and Cetaceans: Field Studies and Conservation of Complex Mammalian Societies, Springer, New York, NY.
- Ford, J.K.B., A.L. Rambeau, R.M. Abernethy, M.D. Boogaards, L.M. Nichol et L.D. Spaven. An assessment of the potential for recovery of humpback whales off the Pacific coast of Canada, document de recherche 2008/0xx du Secrétariat consultatif de consultation scientifique, ministère des Pêches et des Océans, Ottawa, Ontario, 2008.
- Frouin, H., M. Lebeuf, R. Saint-Louis, M.O. Hammill, É. Pelletier et M. Fournier. « Toxic effects of tributyltin and its metabolites on harbour seal (Phoca vitulina) immune cells in vitro », dans Aquatic Toxicology, vol. 90, 2008, p. 243-251.
- Gade, A., Harwood L., Melling H., Stern G., S.H. Ferguson. Sous presse. « Organochlorine contaminants in ringed seals (Phoca hispida) in the western Canadian Arctic: Trends with sea ice », dans Organohalogen Compounds, vol. 70, nº XXX-XXX.
- Gajadhar, A.A., L.B. Forbes et L.N. Measures. Effect of salt water and chemical disinfectants on the sporulation and infectivity to Toxoplasma gondii oocysts. Proceedings of the American Association of Veterinary Parasitologists, 51e Assemblée annuelle, Honolulu, 15-18 juillet 2006.
- Gervaise, C., S. Vallez, Y. Stephan, C. Iaona et Y. Simard. « Passive acoustic tomography: new concepts and applications using marine mammals », dans Journal of the Marine Biological Association of the United Kingdom, vol. 87, 2007, p. 5-10.
- Gervaise, C., S. Vallez, Y. Stephan et Y. Simard. « Robust 2D localization of low-frequency calls in shallow waters using modal propagation modeling », dans Acoustique canadienne, vol. 36, nº 1, 2008, p. 153-159.
- Gosselin, J.-F., M.O. Hammill et V. Lesage. Comparaison des indices d'abondance photographique et visuels des bélugas de l'estuaire du Saint-Laurent en 2003 et 2005, document de recherche 2007/25 du Secrétariat canadien de consultation scientifique, 2007.
- Gregr, E.J., L.M. Nichol, J.C. Watson, J.K.B. Ford et G.M. Ellis. « Estimating carrying capacity for sea otters in British Columbia », dans Journal of Wildlife Management, vol. 72, 2008, p. 382-388.
- Guinet, C., P. Domenici, R. de Stephanis, L. Barrett-Lennard, J.K.B. Ford et P. Verborgh. « Killer whale predation on bluefin tuna: exploring the hypothesis of the endurance-exhaustion technique », dans Marine Ecology Progress Series, vol. 347, 2007, p. 111-119.
- Hammill, M.O. et G.B. Stenson. Abondance des phoques à capuchon dans l'Atlantique Nord-Ouest (1960 - 2005), document de recherche 2006/068 du Secrétariat canadien de consultation scientifique du MPO, 2006,19 p.
- Hammill, M.O. et G.B. Stenson. Période de mise bas chez les phoques à capuchon du Nord-Ouest, document de recherche 2006/069 du Secrétariat canadien de consultation scientifique, 2006.
- Hammill, M.O., J.F. Gosselin et G.B. Stenson. « Changes in abundance of grey seals in the NW Atlantic », pages 99-115, dans T. Haug, M. Hammill et D. Olafsdottir. (sous la dir. de). Grey seals in the North Atlantic and the Baltic, publication scientifique de la NAMMCO, vol. 6, 2007, 227 p.
- Hammill, M.O., J.W. Lawson, G.B. Stenson et D. Lidgard. La production de phoques gris de l'Atlantique Nord Ouest dans le golfe du Saint Laurent et le long de la côte est de la Nouvelle Écosse, document de recherche 2007/084 du Secrétariat canadien de consultation scientifique du MPO, 2007.
- Hammill, M.O., V. Lesage, P. Richard et P. Hall. « A response to ‘sentient beings and wildlife resources: Inuit, beluga whales and management regimes in the Canadian Arctic », dans Human Ecology, vol. 35, nº 5, 2007, p. 639-640.
- Hammill, M.O., L.N. Measures, J.-F. Gosselin et V. Lesage. Absence de rétablissement du béluga de l'estuaire du Saint-Laurent, document de recherche 2007/026 du Secrétariat canadien de consultation scientifique, 2007, 23 p.
- Hammill, M.O. et G.B. Stenson. « Application of the precautionary approach and conservation reference points to management of Atlantic seals », dans ICES Journal of Marine Science, vol. 64, nº 4, 2007, p. 702-706.
- Hammill, M.O., G.B. Stenson, F. Proust, P. Carter et D. McKinnon. « Feeding by grey seals in the Gulf of St. Lawrence and around Newfoundland », pages 135-152, dans T. Haug, M. Hammill et D. Olafsdottir (sous la dir. de). Grey seals in the North Atlantic and the Baltic, publication scientifique de la NAMMCO, vol. 6. 2007, 227 p.
- Harris, L. Évaluation du potentiel de rétablissement de la baleine à bec commune, population du plateau néo-écossais, rapport 2007/011 du Secrétariat canadien de consultation scientifique, 2007.
- Harvey, V., S. D. Côté et M.O. Hammill. « The Ecology Of 3-D Space Use In A Sexually Dimorphic Mammal », dans Ecography, vol. 31, 2008, p. 371-380.
- Harwood, L.A., T.G. Smith et H. Melling. « Assessing the potential effects of near shore hydrocarbon exploration on ringed seals in the Beaufort Sea Region 2003-2006 », dans Environmental Studies Research Funds, vol. 162, 2007, 103 p.
- Harwood, L.A. et P. Sparling. « Lake trout distribution and salinity: an assessment of the relative abundance and distribution of lake trout throughout Husky Lakes, 2001-2004 », dans Mills, K. H., M. Dyck et L. A. Harwood. « Proceedings of the Second North American Lake Trout Symposium 2005, Yellowknife, Northwest Territories » dans Canadian Technical Report of Fisheries and Aquatic Sciences, vol. 2778, 2008, 247 p.
- Harwood, L.A., A. Joynt et S. Moore. Bowhead whale feeding aggregations in the Canadian Beaufort Sea and their role in the mitigation of effects of seismic underwater noise. Document de travail et exposé au Deuxième atelier d'évaluation des renseignements scientifiques sur les impacts des bruits sismiques sur les poissons, les invertébrés et les mammifères marins, 26-28 mars, 2008, Ottawa, Ontario.
- Harwood, L.A., F. Pokiak et J. Walker-Larsen. « Assessment of the subsistence fishery and biological data for Arctic cisco in Tuktoyaktuk Harbour, NT, Canada, 1997-1999 », dans Canadian Management Report of Fisheries and Aquatic Sciences. vol. 2845, 2008, ix + 31 p.
- Haug, T., G.B. Stenson, P.J. Corkeron et K.T. Nilssen. « Estimation of harp seal pup production in the North Atlantic completed: results from surveys in the Greenland Sea in 2002 », dans ICES Journal of Marine Science, vol. 63, nº 1, 2006, p. 95-104.
- Heide-Jorgensen, M.P., K.L. Laidre, M.V. Jensen, L.P. Dueck et L.D. Postma. « Dissolving stock discreteness with satellite tracking: bowhead whales in Baffin Bay », dans Marine Mammal Science, vol. 22, 2006, p. 34-45.
- Heise, K., J. Ford et P. Olesiuk. « Marine mammals and turtles », dans Lucas, B.G., Verrin, S. et R. Brown. (sous la dir. de). « Ecosystem overview: Pacific North Coast Integrated Management Area (PNCIMA) », dans Canadian Technical Report of Fisheries and Aquatic Sciences, vol. 2667, 2007, xii + 104 p.
- Heise, K., L.G. Barrett-Lennard et J.K.B. Ford. Update COSEWIC Status Report on Killer Whales Orcinus orca, préparé pour le Comité sur la situation des espèces en péril au Canada en 2008.
- Hickie, B.E., P.S. Ross, R.W. Macdonald et J.K.B. Ford. « Killer whales (Orcinus orca) face protracted health risks associated with lifetime exposure to PCBs », dans Environmental Science and Technology, vol. 41, nº 18, 2007, p. 6613-6619.
- Higdon, J. État des connaissances sur les épaulards (Orcinus orca) dans les eaux canadiennes de l'Arctique, document de recherche 2007/048 du Secrétariat canadien de consultation scientifique, 2007.
- Higdon J.W., O.R.P Bininda-Emonds, R.M.D. Beck et S.H. Ferguson. « Phylogeny and divergence of the pinnipeds assessed using a multigene dataset », dans BMC Evolutionary Biology, vol. 7, 2007, p. 216.
- Higdon, J.W., et S.H. Ferguson. Sous presse. « Sea ice declines causing punctuated change as observed with killer whale (Orcinus orca) sightings in the Hudson Bay region over the past century », dans Ecological Applications.
- Higdon, J.W., O.R.P. Bininda-Emonds, R.M.D. Beck, et S.H Ferguson. « Correction: Phylogeny and divergence of the pinnipeds (Carnivora: Mammalia) assessed using a multigene dataset », dans BMC Evolutionary Biology, vol. 8, 2008, p. 216.
- Koen-Alonso, M. et G. Stenson. « Keystone species: definitions and related concepts in an applied context », p. 9-20 dans Information scientifique de base pour le choix des critères de détermination des espèces et des propriétés des communautés d'importance écologique, J. Rice (sous la dir. de), document de recherche 2006/089 du Secrétariat canadien de consultation scientifique du MPO, 2006, 82 p.
- Koschinski, S., B.M. Culik, E.A. Trippel et L. Ginzkey. « Behavioral reactions of free-ranging harbor porpoises encountering standard nylon and BaSO4 mesh gillnets and warning sound », dans Marine Ecology Progress Series, vol. 313, 2006, p. 285-294.
- Krahn, M.M., M.B. Hanson, R.W. Baird, D.G. Burrows, C.K. Emmons, J.K.B. Ford, L.L. Jones, D.P. Noren, P.S.Ross, G.S. Schorr, T.K. Collier et R.H. Boyer. « Persistent organic pollutants and stable isotopes in biopsy samples (2004/2006) from Southern Resident killer whales », dans Marine Pollution Bulletin, vol. 54, nº 12, 2007, p. 1903-1911.
- Laidre, K.L., I. Stirling, L.F. Lowry, Ø. Wiig, M.P. Heide-JØrgensen et S.H. Ferguson. « Quantifying the sensitivity of Arctic marine mammals to climate-induced habitat change », dans Ecological Applications, vol.18, 2008, p. S97-S125.
- Lambourn, D.M., S.J. Jeffries, M.M. Lance, P.F. Olesiuk, S.A. Raverty et A.W. Trites. Blood analysis on free ranging Steller sea lions (Eumetopias jubatus), International Association for Aquatic Animal Medicine Conference, 6-10 mai 2006. Nassau, Bahamas.
- Lapierre, J.L., J.F. Schreer, J.M. Burns et M.O. Hammill. « Effect of Diazepam on heart and respiratory rates of Harbour seal pups following intravenous injection », dans Marine Mammal Science, vol. 23, nº 1, 2007, p. 209-217.
- Lawson, J., M. Hammill et G. Stenson. Caractéristiques pour le rétablissement: Béluga, document de recherche 2006/075 du Secrétariat canadien de consultation scientifique du MPO, 2006.
- Lebeuf, M., M. Noël, S. Trottier et L.N. Measures. « Temporal trends (1987-2002) of persistent, bioaccumulative and toxic (PBT) chemicals in beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary, Canada », dans Science of the Total Environment, vol. 383, 2007, p. 216-231.
- Leblanc, E., M. Bahoura et Y. Simard. « Comparison of automatic classification methods for beluga whales vocalizations », dans Journal of the Acoustical Society of America, vol. 123, 2008, p. 3772.
- Ledwell, W., S. Benjamins, J. Lawson et J. Huntington. « The most southerly record of a stranded bowhead whale, Balaena mysticetus, from the western North Atlantic Ocean », dans Arctic, vol. 60, nº 1, 2007, p. 17-22.
- Lesage, V., J. Keays, S. Turgeon et S. Hurtubise. « Bycatch of harbour porpoises (Phocoena phocoena) in the gillnet fishery of the Estuary and Gulf of St Lawrence, 2000--2002 », dans Journal of Cetacean Research and Management, vol. 8, nº 1, 2006, p. 67-78.
- Lesage, V., J.-F. Gosselin, M.O. Hammill, M.C.S. Kingsley et J. Lawson. Zones d'importance écologique et biologique (ZIEB) pour l'estuaire et le golfe du Saint-Laurent - une perspective des mammifères marins, document de recherche 2007/046 du Secrétariat canadien de consultation scientifique, 2007.
- Lewis, A.E., Hammill, M.O., Power, M., Doidge, D.W. et V. Lesage. Sous presse. « A comparison of eastern Hudson Bay beluga (Delphinapterus leucas) movement and aggregation patterns using satellite telemetry and traditional ecological knowledge », dans Arctic, 62 (1): 13-24.
- Lidgard, D. et W.D. Bowen. Interactions comportementales entre les prédateurs et leurs proies dans les écosystèmes marins, document de recherche 2008/032 du Secrétariat canadien de consultation scientifique, 2008.
- Lidgard, D.C., D.J. Boness, W.D. Bowen et J.I. McMillan. « The implications of stress on male mating behavior and success in a sexually dimorphic polygynous mammal, the grey seal », dans Hormones and Behaviour, vol. 53, 2008, p. 241-248.
- Lilly, G., K. Wieland, B. J. Rothschild, S. Sundby, K.F. Drinkwater, K. Brander, G. Ottersen, J.E. Carscadden, G.B. Stenson, G.A. Chouinard, D.P. Swain, N. Daan, K. Enberg, M.O. Hammill, A. Rosing-Asvid, H. Svedäng et A. Vázquez. « Decline and Recovery of Atlantic Cod (Gadus morhua) Stocks throughout the North Atlantic », 2008, dans G.H. Kruse, K. Drinkwater, J.N. Jianelli, J.S. Link, D.L. Stram, V. Wespestad et D. Woodby (sous la dir. de). Resiliency of Gadid stocks to fishing and climate change, AK-SG-08-01, 2008.
- Loseto, L., P. Richard, J. Orr, G. Stern, et S.H. Ferguson. « Intrapopulation habitat selection by beluga whales in the eastern Beaufort Sea », dans Revue canadienne de zoologie, vol. 84, 2006, p. 1743-1751.
- Loseto, L.L., G.A. Stern, et S.H. Ferguson. « Size and Biomagnification: How habitat selection explains beluga mercury levels », dans Environmental Science and Technology, vol. 42, p. 3982-3988, 2008, 10.1021/es7024388.
- Loseto, L.L., G.A. Stern, D. Deibel, T.L. Connelly, A. Prokopowicz, D.R.S. Lean, L. Fortier et S.H. Ferguson. « Linking mercury exposure to habitat and feeding behaviour in Beaufort Sea beluga whales », dans Journal of Marine Systems, vol. 74, 2008, p. 1012-1024.
- Luque, S. P., J. Higdon et S. H. Ferguson. « Dentine deposition rates in beluga (Delphinapterus leucas): an analysis of the evidence », dans Journal of Aquatic Mammals, vol. 33, 2007, p. 241-245.
- Matkin, C.O., E.L. Saulitis, G.M. Ellis, P. Olesiuk, et S.D. Rice. « Ongoing population-level impacts on killer whales Orcinus orca following the ‘Exxon Valdez' oil spill in Prince William Sound, Alaska », dans Marine Ecology Progress Series, vol. 356, 2008, p. 269-281.
- Measures, L.. Compte rendu de l'atelier sur le béluga de l'estuaire du Saint-Laurent - Revue du programme des carcasses, compte rendu 2007/005 du Secrétariat canadien de consultation scientifique, 2007.
- Miller, E. H., H.-C. Sing, V.D. Moulton, G.W. Miller, J.K. Finley et G.B. Stenson. « Variation and integration of the simple mandibular postcanine dentition in two species of phocid seals », dans Journal of Mammalogy, vol. 88, 2007, p. 1325-1334.
- Mills, K.H., M. Dyck et L.A. Harwood. « Proceedings of the Second North American Lake Trout Symposium 2005, Yellowknife, Northwest Territories », dans Canadian Technical Report of Fisheries and Aquatic Sciences, vol. 2778, 2008, 247 p.
- Mooney T.A., W.W.L. Au, P.E. Nachtigall et E.A. Trippel. « Acoustic and stiffness properties of gillnets as they rebate to small cetacean bycatch », dans ICES Journal of Marine Science, vol. 64, nº 7, 2007, p. 1324-1332.
- Morissete, L., M.O. Hammill et C. Savenkoff. « The trophic role of marine mammals in the northern Gulf of St. Lawrence », dans Marine Mammal Science, vol. 22, nº 1, 2006, p. 74-103.
- Mos, L., B. Morsey, S.J. Jeffries, M.B. Yunker, S. Raverty, S. De Guise et P.S. Ross. « Chemical and biological pollution contribute to the immunological profiles of free-ranging harbor seals », dans Environmental Toxicology and Chemistry, vol. 25, nº 12, 2006, p. 3110-3117.
- Mos, L., M. Tabuchi, N. Dangerfield, S.J. Jeffries, B.F. Koop et P.S. Ross. « Contaminant-associated disruption of vitamin A and its receptor (retinoic acid receptor alpha) in free-ranging harbour seals (Phoca vitulina) », dans Aquatic Toxicology, vol. 81, nº 3, 2007, p. 319-328.
- Nichol, L.M. Évaluation du potentiel de rétablissement de la loutre de mer (Enhydra lutris) au Canada, document de recherche 2007/034 du Secrétariat canadien de consultation scientifique, 2007.
- Nielsen, O., G. Smith, H. Weingartl, S. Lair et L. Measures. « Use of a SLAM transfected Vero cell line to isolate and characterize marine mammal morbilliviruses using an experimental ferret model », dans Journal of Wildlife Diseases, vol. 44, 2008, p. 600-611.
- Noren, S. R., D.J. Boness, S.J. Iverson, J. McMillan et W.D. Bowen. « Body Condition at Weaning Affects the Duration of the Postweaning Fast in Gray Seal Pups (Halichoerus grypus) », dans Physiological & Biochemical Zoology, vol. 81, 2008, p. 269-277.
- Olesiuk, P.F. Évaluation préliminaire du potentiel de rétablissement des otaries à fourrure du Nord (Callorhinus ursinus) de Colombie-Britannique, document de recherche 2007/006 du Secrétariat canadien de consultation scientifique, 2007.
- Olesiuk, P.F. Abondance de l'otarie de Steller (Eumetopias jubatus) en Colombie-Britannique, document de recherche 2008/063 du Secrétariat canadien de consultation scientifique, 2008.
- Olesiuk, P.F., D.G. Calkins, K.W. Pitcher, W.L. Perryman, C. Stinchcomb et M. Lynn. Évaluation des dénombrements de petits de l'otarie de Steller (Eumetopias jubatus) à partir d'images 35 mm prises à angle oblique, document de recherche 2008/064 du Secrétariat canadien de consultation scientifique, 2008.
- Pang, D.S.J., Y. Rondenay, L.N. Measures et S. Lair. « The effects of two dosages of midazolam on short-duration anesthesia in the harp seal », dans Journal of Zoo and Wildlife Medicine, vol. 37, 2006, p. 27-32.
- Pang, D.S.J., Y. Rondenay, E. Troncy, L.N. Measures et S. Lair. « Use of end-tidal partial pressure of carbon dioxide to predict arterial partial pressure of carbon dioxide in harp seals during isoflurance-induced anesthesia », dans American Journal of Veterinary Research, vol. 67, 2006, p. 1131-1135.
- Park, C.W., Y. Simard, M. Bahoura, J. Rouat et M. Sirois. The remote monitoring system for the buoy to localize the whale by Labview, compte rendu d'OCEANS'06 MTS/IEEE – Boston, IEEE, nº de cat. 06CH37757C, ISBN: 1-4244-0115-1, IEEE, Piscataway, NJ, USA, 2006.
- Parsons, K.M., K.C. Balcomb III, J.K.B. Ford et J.W. Durban. Sous presse. « The social dynamics of southern resident killer whales and conservation implications for this endangered population », dans Animal Behaviour.
- Pitcher, K.W., P.F. Olesiuk, R.F. Brown, M.S. Lowry, S.J. Jeffries, J.L. Sease, W.L. Perryman, C.E. Stinchcomb et L.F. Lowry. « Abundance and distribution of the eastern North Pacific Steller sea lion (Eumetopias jubatus) population », dans Fishery Bulletin, vol. 150, nº 1, 2007, p. 102-115.
- Pitman, R. L., H. Fearnbach, R. LeDuc, J.W. Gilpatrick Jr., J.K.B. Ford et L.T. Balance. « Killer whales prey on a blue whale calf on the Costa Rica Dome: genetics, morphometrics, vocalizations and composition of the group », dans Journal of Cetacean Research and Management, vol. 9, 2007, p. 151-157.
- Raverty, S., B. Hanson, D. Lamborn, J. Gaydos, M. Haulena, J.K.B. Ford, J. Calambokidis et P.S. Ross. An overview of emerging infectious and noninfectious diseases in the Northeastern Pacific Ocean, document de travail, Comité scientifique de la Commission baleinière internationale, SC/59/DW, 59e assemblée annuelle, 7-18 mai 2007, Anchorage, AK, 2007.
- Riesch, R., J.K.B. Ford et F. Thomsen. « Whistle sequences in wild killer whales (Orcinus orca) », dans Journal of the Acoustical Society of America, vol. 124, 2008, p. 1822-1829.
- Robbins, J., J. Barlow, A.M. Burdin, J. Calambokidis, P.J. Clapham, J.K.B. Ford, C.M. Gabriele, R. LeDuc, D. Mattila, T.J. Quinn II, L. Rojas-Bracho, J.M. Straley, J. Úrban, P. Wade, D. Weller, B.H. Witteveen, K. Wynne et M. Yamaguchi. Minimum estimates of humpback whale entanglement frequency in the North Pacific Ocean based on scar evidence, document de travail SC/59/BC, Comité scientifique de la Commission baleinière internationale, Anchorage, AK, mai 2007.
- Ross, P.S. « Fireproof killer whales: Flame retardant chemicals and the conservation imperative in the charismatic icon of British Columbia », dans Journal canadien des sciences halieutiques et aquatiques, vol. 63, 2006, p. 224-234.
- Roy, N., Y. Simard et J. Rouat. « Performance of three acoustical methods for localizing whales in the Saguenay-St. Lawrence Marine Park », dans Acoustique canadienne, vol. 36, nº 1, 2008, p. 160-164.
- Roy, N., Y. Simard, C. Bédard et E. Leblanc. « Habitat occupation time-series of St. Lawrence belugas from passive acoustic monitoring », dans Journal of the Acoustical Society of America. vol. 123, 2008, p. 3771-3772.
- Savenkoff, C., M. Castonguay, D. Chabot, A. Fréchet, M.O. Hammill et L. Morissette. « Main prey and predators and estimates of mortality of Atlantic cod (Gadus morhua) in the northern Gulf of St. Lawrence during the mid-1980s, mid-1990s, and early 2000s », dans Canadian Technical Report of Fisheries and Aquatic Sciences, vol. 2666, 2006, 40 p.
- Savenkoff, C., D.P. Swain, J.M. Hanson, M. Castonguay, M.O. Hammill, H. Bourdages, L. Morissette et D. Chabot. « Effects of fishing and predation in a heavily exploited ecosystem: Comparing periods before and after the collapse of groundfish in the southern Gulf of St. Lawrence (Canada) », dans Ecological Modelling, vol. 204, 2007, p. 115-208.
- Savenkoff, C., M. Castonguay, D. Chabot , M. O. Hammill , H. Bourdages et L. Morissette. « Changes in the northern Gulf of St. Lawrence ecosystem estimated by inverse modelling: Evidence of a fishery-induced regime shift? », dans Estuarine, Coastal and Shelf Science, vol. 73, 2007, p. 711-724.
- Savenkoff, C., L. Morissette, M. Castonguay, D.P. Swain, M. O. Hammill, D. Chabot et J.M. Hanson. « Interactions between marine mammals and fisheries: Implications for cod recovery », pages 107-151, dans Ecosystem Ecology Research Developments (F. Columbus, sous la dir. de), Nova Science Publishers, Inc., New York, 2008.
- Shelton, P.A., B. Best, A. Cass, C. Cyr, D. Duplisea, J. Gibson, M. Hammill, S. Khwaja, M.A. Koops, K.A. Martin, R. O'Boyle, J.C. Rice, A. Sinclair, K. Smedbol, D.P. Swain, L.A. Vélez-Espino et C.C. Wood. Évaluation du potentiel de rétablissement: projections à long terme, document de recherche 2007/045 du Secrétariat canadien de consultation scientifique, 2007.
- Simard, Y., N. Roy et C. Gervaise. Shipping noise and whales: world's tallest ocean liner vs. largest animal on earth, OCEANS'06 MTS/IEEE – Boston, IEEE, nº de cat. 06CH37757C, ISBN: 1-4244-0115-1, IEEE, Piscataway, NJ, USA, 2006.
- Simard, Y., C. Bédard, X. Mouy et L. Fortier. Passive acoustic detection of bowheads, belugas and seals at Mackenzie Shelf break in Beaufort Sea during fall 2004, réunion scientifique annuelle ArcticNet des RCE du CRSNG, Victoria, C.-B., 12-15 décembre 2006.
- Simard,Y., M. Bahoura, C.W. Park, J. Rouat, M. Sirois, X. Mouy, D. Seebaruth, N. Roy et R. Lepage. Development and experimentation of a satellite buoy network for real-time acoustic localization of whales in the St. Lawrence, compte rendu d'OCEANS'06 MTS/IEEE – Boston, IEEE, Piscataway, NJ, USA, 2006.
- Simard, Y. et A. Stepnowski. « Classification methods and criteria », chapitre 5, dans Anderson et coll. (sous la dir. de). « Acoustic seabed classification of marine physical and biological landscapes », dans Rapport des recherches collectives du CIEM, nº 286, 2007, p. 61-72.
- Simard, Y. et J.T. Anderson. « Terms and acronyms », dans Anderson et coll. (sous la dir. de). « Acoustic seabed classification of marine physical and biological landscapes », dans Rapport des recherches collectives du CIEM, nº 286, 2007, p. 147-151.
- Simard, Y., V. Lesage, J.-F. Gosselin, M. Hammill, N. Roy et C. Cyr. Impacts de la construction d'un port méthanier à Gros-Cacouna sur les mammifères marins, réponse des Sciences 2007/010 du Secrétariat canadien de consultation scientifique du MPO, 2007.
- Simard, Y. Sous presse. « The Saguenay-St. Lawrence Marine Park: oceanographic process at the basis of this unique forage site of Northwest Atlantic whales », dans Rev. Sc. Eau.
- Simard, Y. et L. Fortier. Listening at climate change in ice-covered seas: example with AURAL autonomous hydrophones in the Arctic, compte rendu de l'IEEE Oceans'08. MTS/IEEE - Québec, IEEE, Piscataway, NJ. USA, 2008, 2 p.
- Simard, Y. et N. Roy. « Detection and localization of blue and fin whales from large-aperture autonomous hydrophone arrays: a case study from the St. Lawrence estuary », dans Acoustique canadienne, vol. 36, nº 1, 2008, p. 104-110.
- Simard, Y. N. Roy et C. Gervaise. « Passive acoustic detection and localization of whales: effects of shipping noise in Saguenay-St. Lawrence Marine Park », dans Journal of the Acoustical Society of America, vol. 123, nº 6, 2008, p. 4109-4117.
- Simard, Y., N. Roy et C. Gervaise. « Masking of blue and fin whales low-frequency vocalizations by shipping noise in the Saguenay—St. Lawrence Marine Park », dans Bioacoustics, vol. 17, 2008, p. 183-185.
- Simard, Y., N. Roy, F.-J. Saucier, J. Gagné et S. Giard. « Saguenay fjord entrance whale feeding ground: Acoustic study of sill dynamics and tidal aggregation of forage fish », dans Journal of the Acoustical Society of America, vol. 123, 2008, p. 2992-2993.
- Simard, Y. et Sourisseau, M. Sous presse. « Diel changes in acoustic and catch estimates of krill biomass », dans ICES Journal of Marine Science, XX: XX-XXX.
- Smedbol, R.K. Évaluation du potentiel de rétablissement de la baleine noire (population de l'ouest de l'Atlantique Nord (Eubalaena glacialis) dans les eaux canadiennes, document de recherche 2007/044 du Secrétariat canadien de consultation scientifique, 2007.
- Sourisseau, M., Y. Simard et F.J. Saucier. « Krill aggregation in the St. Lawrence system, and supply of krill to the whale feeding grounds in the estuary from the gulf », dans Marine Ecology Progress Series, vol. 314, 2006, p. 257-270.
- Sourisseau, M., Y. Simard et F.-J. Saucier. « Krill diel vertical migration fine dynamics, nocturnal overturns, and their roles for aggregation in stratified flows », dans Journal canadien des sciences halieutiques et aquatiques, vol. 65, 2008, p. 574-587.
- Steiger, G. H., J. Calambokidis, J.M. Straley, L.M. Herman, S. Cerchio, D.R. Salden, R. Urbán, J.K. Jacobsen, O. von Ziegesar, K.C. Balcomb, C.M. Gabriele, M.E. Dahlheim, S. Uchida, J.K.B. Ford, P. Ladrón de Guevara, M. Yamaguchi et J. Barlow. « Geographic variation in killer whale attacks on humpback whales in the North Pacific: implications for predation pressure », dans Endangered Species Research, vol. 4, 2008, p. 247-256.
- Straley, J., J. Ford, G. Ellis, D. Matkin, V. Deecke et C. Gabriele. Prey preferences of mammal-eating killer whales in southeastern Alaska and British Columbia, document de travail présenté à la réunion spéciale sur les épaulards, Comité scientifique de la Commission baleinière internationale, SC/59/Rep 1, 59e assemblée annuelle, 7-18 mai 2007, Anchorage, AK, 2007.
- Stewart, R.E.A. S. E. Campana, C. M. Jones et B.E. Stewart. « Bomb radiocarbon dating calibrates beluga (Delphinapterus leucas Pallas, 1776) age estimates », dans Revue canadienne de zoologie, vol. 84, nº 12, 2006, p. 1840-1852.
- Stenson, G. Estimating abundance of seals in the North Atlantic: designing and conducting a scientific assessment, p. 47 -51. NAMMCO Conference on User Knowledge and Scientific Knowledge in Management Decision-Making, G. K. Hovelsrud et C. Winsnes (sous la dir. de), Reykjavik, Islande, 4-7 janvier 2003, 2006.
- Stenson, G. B. Mortalité causée par la chasse chez les phoques à capuchon de l'Atlantique Nord-Ouest, document de recherche 2006/066 du Secrétariat canadien de consultation scientifique du MPO, 2006.
- Stenson, G. B. et M.O. Hamill. « Estimating consumption of Atlantic cod (Gadus morhua) by harp seals (Pagophilus groenlandicus) », dans Report of the Workshop on the Decline and Recovery of cod Stocks throughout the North Atlantic, including tropho-dynamic effects (WKDRCS), ICES CM 2006/OCC, vol. 12, 2006, p. 152-154.
- Stenson, G.B., M.O. Hammill, J. Lawson et J. F. Gosselin. Production de nouveaux-nés chez les phoques à capuchon, Cystophora cristata, dans l'Atlantique Nord-Ouest en 2005, document de recherche 2006/067 du Secrétariat canadien de consultation scientifique du MPO, 2006.
- Stenson, G. B. Current research on the impact of pinnipeds on commercial fish stocks in the Northwest Atlantic, NAFO SCR Doc. 07/49, nº de série N5401, 2007.
- Tabuchi, M., A. Veldhoen, N. Dangerfield, S. Jeffries, C.C. Helbing et P.S. Ross. « PCB-related alteration of thyroid hormones and thyroid hormone receptor gene expression in free-ranging Harbor Seals » dans Environmental Health Perspectives, vol. 114, nº 7, 2006, p. 1024-1031.
- Thomas, L, M.O. Hammill et W.D. Bowen. Évaluation de la population de phoques gris de l'Atlantique Nord-Ouest, 1977 – 2007, document de recherche 2007/082 du Secrétariat canadien de consultation scientifique, 2007.
- Tollit, D.J, A.D. Schulze, A.W. Trites, P.F. Olesiuk, S.J. Crockford, T.S. Gelatt, R.R. Ream et K.M. Miller. Sous presse. « Development and application of DNA techniques for validating and improving pinniped diet estimates », dans Ecological Applications.
- Trites, A.W., V.B. Deecke, E.J. Gregr, J.K.B. Ford et P.F. Olesiuk. « Killer whales, whaling, and sequential megafaunal collapse in the North Pacific: A comparative analysis of the dynamics of marine mammals in Alaska and British Columbia following commercial whaling », dans Marine Mammal Science, vol. 23, nº 4, 2007, p. 751-756.
- Trzcinski, M.K., R. Mohn et W.D. Bowen. « Continued decline of the threatened Eastern Scotian Shelf Atlantic cod population: how important is grey seal predation », dans Ecological Applications, vol. 16, 2006, p. 2276-2292.
- Tucker, S., W.D. Bowen et S.J. Iverson. « Dimensions of diet segregation in grey seals Halichoerus grypus through stable isotopes of carbon (d13C) and nitrogen (d15N) », dans Marine Ecology Progress Series, vol. 339, 2007, p. 271-282.
- Tucker, S., W.D. Bowen et S.J. Iverson. « Convergence of diet estimates from fatty acids and stable isotopes, support for long-term integration of natural diets within individual grey seals », dans Marine Ecology Progress Series, vol. 354, 2008, p. 267-276.
- Tucker, S., W.D. Bowen, S.J. Iverson et G.B. Stenson. Sous presse. « Intrinsic and extrinsic sources of variation in the diets of harp and hooded seals revealed by fatty acid profiles », dans Canadian Journal of Zoology.
- Veinott, G. et B. Sjare. « Mercury, cadmium, selenium, and seven other elements in the muscle, renal, and hepatic tissue of harbor seals from Newfoundland and Labrador », dans Bulletin of Environmental Contamination and Toxicology, vol. 77, nº 4, 2006, p. 597-607.
- Wade, P.R., V.N. Burkanov, M.E. Dahlheim, N.A. Friday, L.W. Fritz, T.R. Loughlin, S.A. Mizroch, M.M. Muto, D.W. Rice, L.G. Barrett-Lennard, N.A. Black, A.M. Burdin, J. Calamboikidis, S. Cerchio, J.K.B. Ford, J.K. Jacobsen, C.O. Matkin, D.R. Matkin, A.V. Mehta, R.J. Small, J.M. Straley, G.R. VanBlaricom et P.J. Clapham. « Killer whales and marine mammal trends in the North Pacific – A re-examination of evidence for sequential megafauna collapse and the prey-switching hypothesis », dans Marine Mammal Science, vol. 23, nº 4, 2007, p. 766-802.
- Ward, E.J., K. Parsons, E.E. Holmes, K.C. Balcomb III et J.K.B. Ford. Sous presse. « The role of menopause and reproductive senescence in a long-lived social mammal », dans Frontiers in Zoology.
- Weir, J. N., S. P. Mahoney, B. McLaren et S.H. Ferguson. « Impact of mining on woodland caribou (Rangifer tarandus) in Newfoundland, Canada », dans Wildlife Biology, vol. 13, 2007, p. 66-74.
- Wiig, Ø., E.W. Born, I. Gjertz, C. Lydersen et R.E.A. Stewart. « Historical sex-specific distribution of Atlantic walrus (Odobenus rosmarus rosmarus) in Svalbard assessed by mandible measurements », dans Polar Biology, vol. 31, nº 1, 2007, p. 69-75.
- Wolkers, H., M.O. Hammill et B. van Bavel. « Tissue-specific accumulation and lactational transfer of polychlorinated biphenyls, chlorinated pesticides, and brominated flame retardants in hooded seals from the Gulf of St. Lawrence: Applications for monitoring », dans Environmental Pollution, vol. 142, nº 3, 2006, p. 476-486.
- Wolkers, H., F. Boily, F. van Bavel, M.O. Hammill, J. Fink-Gremmels et R. Primicerio. « Tissue-specific contaminant accumulation and associated effects on hepatic serum analytes and cytochrome P450 enzymes activity in hooded seals (Cystophora cristata) from the Gulf of St. Lawrence: implications for effect studies », dans Archives of Environmental Contamination and Toxicology, DOI 10.1007/s00244-008-9186-6, 2008.
- Zwanenburg, K.C.T., A. Bundy, P. Strain, W.D. Bowen, H. Breeze, S.E. Campana, C. Hannah, E. Head et D. Gordon. « Implications of ecosystem dynamics for the management of the eastern Scotian Shelf », dans Canadian Technical Report of Fisheries and Aquatic Sciences, vol, 2652, nº xiii, 2006, 91 p.
- Date de modification :