
Centre d'expertise sur les mammifères marins : Rapport de recherche scientifique 2009-2011
Table des matières
- Remerciements
- Message du directeur du CEMAM
- 1.0 Dynamique des populations
- 1.1 Croissance des populations de mammifères marins dépendante et indépendante de la densité (Mike Hammill, Don Bowen, Garry Stenson)
- 1.2 Changements dans les taux de reproduction des phoques du Groenland du nord-ouest de l'Atlantique (G.B. Stenson)
- 1.3 Les changements à long terme des indices vitaux des phoques gris sont liés à une réduction du taux de croissance de la population (Don Bowen)
- 1.4 Reproduction des phoques annelés dans la baie d'Hudson (Steve Ferguson, Magaly Chambellant)
- 1.5 Utilisation de la génétique pour acquérir des connaissances sur les groupes de bélugas dans la baie d'Hudson (Mike Hammill, Lianne Postma)
- 2.0 Rôle des mammifères marins dans l'écosystème
- 2.1 Résumé du processus d'évaluation zonale lié aux répercussions des phoques gris sur les populations de poissons dans l'Est du Canada (Don Bowen)
- 2.2 Associations en mer des phoques gris : aperçus fournis par un nouvel enregistreur de données (Don Bowen)
- 2.3 Alimentation spécialisée des épaulards sur le saumon quinnat (John K.B. Ford)
- 2.4 Écologie alimentaire du rorqual bleu dans l'estuaire du Saint-Laurent (Véronique Lesage, Thomas Doniol-Valcroze)
- 2.5 Océanographie de l'aire d'alimentation des baleines à fanons : le piège océanographique (Yvan Simard)
- 2.6 Prédation des épaulards de l'Arctique (Steve Ferguson)
- 3.0 Interactions entre les mammifères marins et les humains
- 4.0 Relations avec les conseils de cogestion
- 5.0 Espèces en péril
- 6.0 Publications de 2009-2011
Remerciements
Cette publication a été produite grâce aux nombreux scientifiques du MPO spécialistes des mammifères marins de partout au Canada qui ont fourni des commentaires et des conseils. Nous remercions le personnel de soutien, les examinateurs et tous ceux qui ont offert une aide précieuse pour la production de cette publication. Nous tenons en outre à remercier Christine Abraham pour toutes les heures de travail consacrées à l'élaboration du rapport.
Nous aimerions remercier plus particulièrement les personnes suivantes qui ont contribué à la rédaction du texte :
- Don Bowen - Chercheur scientifique, Halifax (Nouvelle-Écosse)
- Thomas Doniol-Valcroze - Biologiste, Mont-Joli (Québec)
- Steve Ferguson - Chercheur scientifique, Mont-Joli (Québec)
- John Ford - Research Scientist, Chercheur scientifique, Nanaimo (Colombie-Britannique)
- Mike Hammill - Chercheur scientifique, Mont-Joli (Québec)
- Lois Harwood - Biologiste, Yellowknife (Territoires du Nord-Ouest)
- Michel Lebeuf - Biologiste, Mont-Joli (Québec)
- Véronique Lesage - Chercheur scientifique, Mont-Joli (Québec)
- Lianne Postma - Biologiste, Winnipeg (Manitoba)
- Peter Ross - Chercheur scientifique, Sidney (Colombie-Britannique)
- Yvan Simard - Chercheur scientifique, Mont-Joli (Québec)
- Sjare Research - Chercheur scientifique, St John's (Terre-Neuve-et-Labrador)
- Garry Stenson - Chercheur scientifique, St John's (Terre-Neuve-et-Labrador)
Message du directeur du CEMAM
Le Centre d'expertise sur les mammifères marins (CEMAM) de Pêches et Océans Canada (le MPO) est un centre virtuel qui comprend une quarantaine de professionnels et d'employés de soutien situés dans sept laboratoires dans tout le pays. La recherche effectuée par le CEMAM est variée; elle comprend les niveaux de contaminants et leurs effets, la structure et la dynamique de la population, l'écologie alimentaire, l'utilisation et les besoins liés à l'habitat, les interactions avec les pêches et les répercussions environnementales du développement. De nombreuses espèces de mammifères marins du Canada n'ont montré que quelques signes de rétablissement suite à leur surexploitation antérieure; la chasse exerce une pression sur des espèces du Nord alors que d'autres espèces se sont complètement rétablies et ne connaissent aucune menace à l'heure actuelle.
Un défi majeur de nos activités est de fournir des conseils scientifiques afin d'informer les politiques et les programmes sur les espèces en péril aussi bien que sur les espèces abondantes. Le ralentissement économique mondial a entraîné de nombreux gouvernements à réexaminer la prestation de leurs programmes. Cette situation aura des répercussions sur nos recherches et sur la façon dont nous offrons des conseils. Par ailleurs, les enjeux émergents doivent faire l'objet d'une attention particulière. Le changement climatique, même s'il n'est pas nouveau, devient de plus en plus évident. Par exemple, le déclin de la couverture de glace dans l'Atlantique limitera probablement la répartition des phoques du Groenland dans le sud de leur aire de reproduction, et favorisera possiblement l'expansion vers le nord des autres espèces comme les phoques gris. Cela pourrait alors augmenter les tensions avec les pêcheurs au sujet des répercussions potentielles des phoques sur la pêche commerciale. On s'attend à ce que le déclin des glaces de l'Arctique mène à une augmentation du trafic maritime afin de fournir un accès à de nouvelles zones de développement commercial. Parallèlement, il réduira considérablement l'habitat de certaines espèces de mammifères marins, tout en créant de nouveaux habitats pour des espèces anciennement tempérées qui pourraient se déplacer vers le nord. Au large des côtes est et ouest du Canada, l'exploitation extracôtière et la navigation maritime accrue, en particulier vers l'Asie, peuvent également avoir une incidence sur les mammifères marins. Par conséquent, une meilleure compréhension du rôle des mammifères marins dans les écosystèmes marins, leurs besoins en matière d'habitat et les répercussions du bruit ainsi que les effets cumulatifs de l'exploitation constituent seulement quelques problèmes qui devront faire l'objet d'une recherche approfondie et qui nécessiteront des conseils de gestion dans les années à venir.
Les progrès technologiques ainsi que l'élaboration de bases de données multidisciplinaires et de méthodes analytiques plus puissantes ont eu des répercussions positives majeures sur la recherche liée aux mammifères marins. Grâce à l'élaboration de nouveaux capteurs, les mammifères marins peuvent maintenant transmettre des renseignements océanographiques aux satellites à partir de zones éloignées, difficiles à échantillonner par bateau. Ces données sont utilisées par les océanographes afin d'améliorer l'exactitude des modèles de circulation océanique. L'utilisation d'émetteurs et de récepteurs acoustiques miniaturisés sur les mammifères marins et leurs proies fournit de nouvelles manières d'étudier comment les mammifères marins s'associent en mer ainsi que les interactions prédateur-proie. L'échantillonnage aux fins de biopsie des grandes baleines peut maintenant être utilisé afin de fournir des renseignements sur les niveaux de stress et l'état reproducteur en plus des données sur le régime alimentaire, les contaminants et la génétique. De nouvelles analyses comprenant de multiples bases de données ont amélioré notre compréhension de l'habitat essentiel et le potentiel d'expansion des aires de distribution, p. ex., les loutres de mer, afin de mieux comprendre les interactions prédateur-proie, de déterminer les zones d'importance écologique et biologique.
Les bénéfices des travaux du CEMAM ne peuvent être pleinement réalisés qu'en les rendant accessibles au plus grand nombre. En plus des ouvrages scientifiques, les résultats de nos travaux sont disponibles sur le site Web du Secrétariat canadien des avis scientifiques (SCAS), ainsi que sur notre propre site Web, par l'entremise d'articles populaires, d'entrevues, de conférences et d'avis par courriel au sujet des nouvelles publications.
Ce rapport ne constitue qu'un échantillon de certaines des activités entreprises par les chercheurs du CEMAM. Il offre un résumé des résultats obtenus sur plusieurs années. Nous avons tenté de fournir une coupe transversale d'activités qui mettent en évidence de nouvelles approches, de nouveaux résultats ainsi que des descriptions plus générales des phénomènes qui ont une incidence sur les populations de mammifères marins au Canada.
Mike Hammill
1.0 Dynamique des populations
1.1 Croissance des populations de mammifères marins dépendante et indépendante de la densité (Mike Hammill, Don Bowen, Garry Stenson)
Les effectifs des populations ont naturellement tendance à changer au fil du temps sous l'influence de facteurs comme la concurrence pour les ressources (p. ex., la nourriture), la prédation, l'immigration, l'émigration et la variabilité environnementale. Un problème qui fait souvent l'objet de discussion est le fait de déterminer si ces changements dans les effectifs constituent le résultat de processus qui dépendent ou non de la densité (figure 1). Les facteurs dépendants de la densité ont une incidence sur les effectifs dont l'ampleur dépend de la taille de la population tandis que les facteurs indépendants de la densité ne sont pas liés à la taille de la population. Les processus limitant les effectifs indépendamment de la densité sont souvent causés par des changements environnementaux comme une fonte soudaine des glaces, qui peut avoir des répercussions catastrophiques pour les espèces de phoques qui ont besoin de la glace comme plateforme pour donner naissance à leurs petits et les élever, ce qui mène à une mortalité élevée des jeunes phoques. El Niño constitue un autre exemple, dans le cadre duquel un changement soudain de la température de l'océan a mené à des pénuries alimentaires à grande échelle, entraînant des famines.
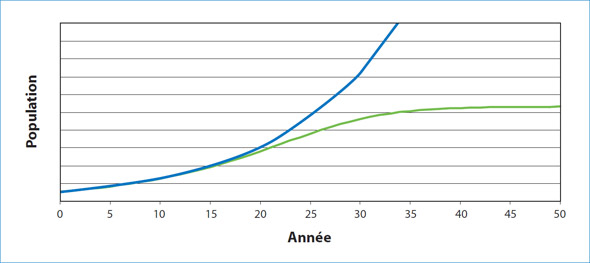
Figure 1. Lorsque la croissance de la population est indépendante de la densité, les ressources ne sont pas limitées et, en théorie, la population augmente de façon continue, souvent de façon exponentielle, jusqu'à ce qu'un événement se produise. Lorsque la croissance de la population est dépendante de la densité, une croissance rapide peut avoir lieu lorsque la population est très petite; toutefois, à un moment donné, la nourriture ou une autre ressource devient limitée et la croissance de la population ralentit et se stabilise à un niveau qu'on appelle la capacité de charge du milieu.
Lorsqu'il y a dépendance à la densité, certaines ressources, habituellement la nourriture (ou l'espace disponible aux fins de reproduction), deviennent de plus en plus limitées; par conséquent, la quantité de nourriture disponible par animal diminue. Sous un certain seuil, cette réduction diminue la condition corporelle, le taux de croissance des individus, le succès reproducteur et en fin de compte la survie. Les effets cette dépendance à la densité ne s'expriment pas uniformément dans la population. Souvent, le premier changement est une augmentation du taux de mortalité des jeunes phoques, surtout au cours de la première année de leur vie, car ils doivent non seulement apprendre à trouver et à capturer de la nourriture, mais doivent aussi consommer plus d'énergie par unité de masse corporelle afin de répondre aux exigences métaboliques et assurer leur croissance. Le taux de survie réduit des jeunes phoques de l'année est souvent associé à une diminution du taux de natalité, car les femelles adultes ont de la difficulté à satisfaire leurs exigences énergétiques de base en plus des coûts supplémentaires de la grossesse et de l'allaitement. Si les conditions deviennent assez graves, la survie des adultes diminue également en réponse au manque de nourriture. Bien que ce modèle général suggère une série de réponses séquentielles, un certain chevauchement aura probablement lieu parmi les changements des indices vitaux pour différentes catégories d'âge et de sexe dans la population.
Lorsque le nombre de naissances dans une population est égal au nombre de décès, la taille de la population se stabilise. Une population stable est considérée comme étant à la capacité de charge du milieu, « K ». Même si l'on tend à la considérer comme étant une capacité constante, K peut varier au fil du temps en réponse à la variabilité environnementale à plus long terme. Par exemple, l'augmentation d'une ressource peut contribuer à améliorer les conditions environnementales, ce qui entraîne une capacité K plus élevée, laquelle a une incidence sur la taille de la population; à l'opposé, des ressources moins élevées peuvent entraîner une diminution de la capacité K.
Il est difficile de détecter lorsqu'une population atteint la capacité de charge ou est en voie de l'atteindre, car certaines fluctuations de la population à court terme sont à prévoir en raison des conditions environnementales variant d'une année à l'autre. De plus, dans le cas des espèces dont nous nous occupons, la taille de la population n'est pas mesurée tous les ans, et chaque estimation de la taille de la population comprend une marge d'erreur. Néanmoins, comme de nombreuses populations de mammifères marins ont été considérablement réduites de façon historique par la chasse, un grand nombre de ces espèces ont augmenté et elles continuent d'augmenter vers la capacité K. Les phoques communs et les baleines grises de la côte ouest du Canada constituent des exemples de populations qui semblent avoir atteint la capacité K ou en être près, tandis que les phoques gris de l'île de Sable présentent des signes de changements dans la croissance de leur population qui sont liés à la densité. Malheureusement, nous n'avons pas de séries à long terme d'estimations d'abondance pour la plupart des populations de mammifères marins; il est donc impossible de déterminer si une population est près de la capacité K ou l'a atteint. De plus, certaines populations sont toujours chassées et il est encore plus difficile de déterminer quelle est la capacité K dans ces situations.
S'il n'y a pas suffisamment de données de relevé pour estimer la capacité K,une autre approche pourrait contribuer à fournir un aperçu de la valeur de la capacité K.
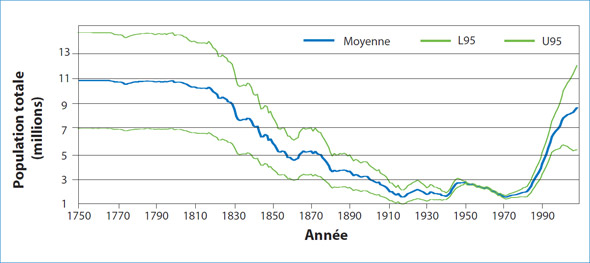
Figure 2: Reconstruction de la population de phoques du Groenland du nord-ouest de l'Atlantique à l'aide de renseignements sur la taille actuelle de la population et de données de chasse datant de la fin des années 1700.
La population de phoques du Groenland du nord-ouest de l'Atlantique a été considérablement réduite en raison de 300 années de chasse. Depuis le début des années 1970, la gestion des quotas a permis à la population d'augmenter et au cours de cette période, nous avons des renseignements sur sa tendance pour une période relativement longue. La population, qui était de moins de deux millions d'animaux au début des années 1970, est passée à près de 8,3 millions d'animaux en 2008; il s'agit de la population la plus importante observée pendant plus de six décennies. Au cours des 30 dernières années, la condition corporelle, la croissance et les taux de reproduction par âge des phoques du Groenland ont montré une tendance au déclin, ce qui suggère que des facteurs dépendants de la densité ont une incidence sur les dynamiques de cette population. Toutefois, des taux de grossesse supérieurs à la moyenne ont entraîné la naissance d'environ 1,6 million de bébés phoques en 2008, ce qui constitue le taux le plus élevé depuis les années 1950. En raison de ces taux de grossesse très variables, il peut être difficile de déterminer si la population est près de sa capacité de charge. Une approche d'estimation de la capacité K serait de tenter de retracer l'histoire de la population jusqu'à une période antérieure au début de la chasse. Les phoques du Groenland font l'objet d'une chasse commerciale depuis le début du 18e siècle. En fait, les chasses aux phoques les plus importantes ont eu lieu au cours de la période des navires de chasse en bois au début des années 1800. Il est très difficile de retracer la population antérieure à la chasse à l'aide des renseignements sur le nombre d'animaux qui ont été pris au fil du temps. Pour conclure que cela reflète la capacité de charge actuelle, il faut supposer que les conditions écologiques qui prévalaient au cours des années 1800 étaient semblables aux conditions environnementales actuelles. Toutefois, lorsque des données de prises historiques ont été inclues dans un modèle et que l'ancienne taille de la population a été retracée, on a pu estimer que la population s'était maintenue à une moyenne de 11 millions d'animaux au début des années 1800. Malheureusement, cette valeur est très incertaine et elle varie possiblement de 7 à 15 millions d'animaux; elle fournit toutefois une indication de la taille maximale possible de cette population (figure 2).
Il est important de comprendre comment les animaux réagissent aux changements de leur abondance. De nombreuses populations de mammifères marins ont été considérablement réduites, et ne comptent aujourd'hui qu'une fraction de leur taille de population vierge. Certaines populations sont maintenant en rétablissement et se rapprochent de leurs abondances historiques. Lorsque les populations sont peu élevées, il y a très peu de différence entre la croissance indépendante de la densité et la croissance dépendante de la densité; dans certains cas, cela a mené à des déclarations affirmant que les populations « explosent » et qu'elles sont hors de contrôle. À un moment donné, les effectifs deviennent toutefois limités par la nourriture, l'espace ou d'autres facteurs, et les facteurs qui dépendent de la densité commencent à limiter la population, ce qui ralentit sa croissance et mène à une stabilisation éventuelle. La compréhension de la nature des changements liés à la mortalité et à la reproduction est essentielle pour l'estimation de l'abondance actuelle et la prévision de la croissance future de la population. La proximité d'une population de la capacité K peut être utilisée en tant qu'indice de sa santé. Une population abondante est plus susceptible de résister aux perturbations soudaines ou aux événements catastrophiques qu'une population plus petite. Les populations saines peuvent également remplir leur rôle écologique traditionnel; pour de nombreux mammifères marins, il s'agit de stabiliser la structure de l'écosystème marin.
1.2 Changements dans les taux de reproduction des phoques du Groenland du nord-ouest de l'Atlantique (G.B. Stenson)
La compréhension des taux de reproduction est essentielle pour comprendre la dynamique des populations de mammifères marins. Comme il est impossible d'étudier la population au complet, nous estimons l'abondance des phoques du Groenland, des phoques à capuchon et des phoques gris en étudiant les petits et en utilisant un modèle de population afin d'extrapoler ces estimations à la taille de la population totale. Par conséquent, la fiabilité de nos estimations de la population dépend de la précision des estimations annuelles du taux de grossesse selon l'âge, d'autant plus que les taux fluctuent en raison de la variation de la taille de l'effectif de la population et des conditions environnementales. Toutefois, le suivi de ces changements est difficile pour la plupart des espèces, car il faut prendre de nombreuses mesures au cours de longues périodes. Depuis les années 1950, des systèmes reproducteurs femelles ont été prélevés sur des phoques du Groenland des eaux de Terre-Neuve et du sud du Labrador; un programme plus systématique entamé dans les années 1980 est toujours en cours aujourd'hui. À l'aide de ces données, les estimations annuelles des taux de grossesse en fin de gestation, de la fécondité (la proportion de femelles adultes qui donnent naissance) et de l'âge moyen de la maturité sexuelle des phoques du Groenland du nord-ouest de l'Atlantique ont été établies.
En général, les taux de grossesse des phoques du Groenland femelles de trois ans étaient très faibles (moins de 10 %); le nombre de femelles enceintes n'est pas élevé. Parmi les phoques de quatre et de cinq ans, les taux de reproduction ont augmenté au cours des années 1970, mais ils ont diminué vers le milieu des années 1980 et sont passés à des niveaux semblables ou inférieurs à ceux des années 1960. Les taux de grossesse pour les femelles de six ans étaient faibles (moins de 67 %) depuis le milieu des années 1990 comparativement aux années précédentes lorsque les taux étaient d'environ 80 %. Parmi les phoques les plus âgés, les taux de grossesse étaient élevés (de 80 à 90 %) jusqu'au milieu des années 1980, mais ils ont ensuite diminué. Depuis, les taux de grossesse ont varié de façon considérable, mais leur moyenne se situe près de 60 %. Depuis 2008, les taux de grossesse ont considérablement diminué; ils sont passés à moins de 30 %. La proportion de femelles adultes qui donnent naissance (le taux de fécondité) suit une tendance semblable; elle est restée relativement élevée jusqu'au milieu des années 1980 puis elle a ensuite diminué (figure 3). Les taux de fécondité récents ont été très variables, les années de faibles taux de grossesse étant associées à des niveaux élevés d'avortements en fin de gestation.

Phoque du Groenland femelle et son petit
Photo : MPO de Terre-Neuve-et-Labrador
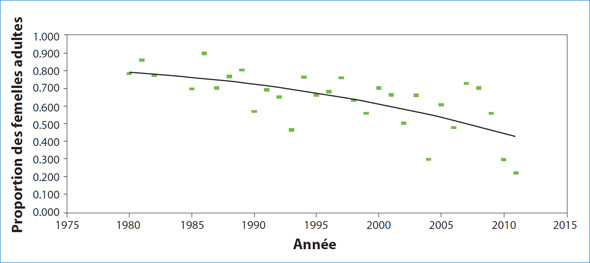
Figure 3: Estimations annuelles de la proportion de phoques du Groenland femelles adultes qui donnent naissance, de 1980 à 2011, en fonction de l'examen des systèmes reproducteurs prélevés au nord-est de Terre-Neuve et au sud du Labrador du mois de décembre au mois de février.

Phoque du Groenland femelle et son petit
Photo : MPO de Terre-Neuve-et-Labrador
L'âge de maturité sexuelle des phoques du Groenland femelles a aussi varié au cours des 50 dernières années. De 1954 à 1976, l'âge moyen de maturité sexuelle était de 5,3 ans. Toutefois, de 1978 à 1987, l'âge moyen de maturité sexuelle des phoques du Groenland femelles est passé à 4,6 ans. En 1990, il a augmenté de près d'une année et au cours de la décennie, l'âge moyen de maturité sexuelle est demeuré relativement stable à 5,6 ans. Sauf en 2000, l'âge moyen de maturité sexuelle a augmenté au début des années 2000 et il a atteint un maximum sans précédent de 6,1 ans en 2005-2006.
Bien que la tendance générale des changements observés corresponde à une réponse de dépendance à la densité de la population, des changements dramatiques dans l'écosystème du nord-ouest de l'Atlantique ont également eu lieu au même moment. Un certain nombre de facteurs physiques et biologiques ont été examinés afin de déterminer ceux qui ont pu avoir une influence sur les taux de grossesse des phoques du Groenland. La diminution générale de la fécondité moyenne semble être un effet de l'augmentation de l'effectif de la population. La variabilité interannuelle est toutefois mieux expliquée par des changements liés aux taux d'avortement en fin de gestation. Les taux d'avortement quant à eux semblent être influencés par les conditions de glace et la disponibilité de la nourriture en début d'hiver, comme l'indiquent les relevés d'abondance de capelans durant les automnes précédents. Cela suggère que la dynamique de population de phoques du Groenland dans le nord-ouest de l'Atlantique est influencée par une interaction complexe de facteurs internes et externes qui comprennent des facteurs physiques et biologiques.
1.3 Les changements à long terme des indices vitaux des phoques gris sont liés à une réduction du taux de croissance de la population (Don Bowen)
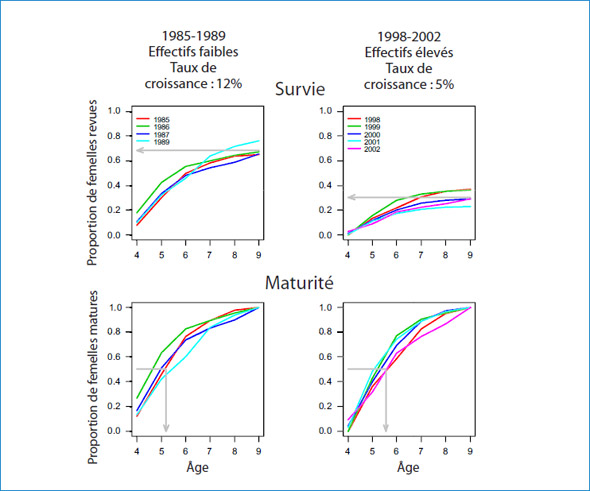
Figure 4: Proportion cumulative des phoques femelles marquées revues sur l'île de Sable entre les âges de 4 et 9 ans (survie, première rangée) et proportion cumulative des femelles revues enceintes ou avec un petit (maturité, deuxième rangée), pour chaque cohorte. Les cohortes marquées de 1985 à 1989, alors que les effectifs étaient faibles et présentaient un taux de croissance de la population élevé, sont situées dans la première colonne, et les cohortes marquées de 1998 à 2002, alors que les effectifs étaient plus élevés et que le taux de croissance était passé de 5 à 4 %, se situent dans la deuxième colonne.
Les populations qui font face à des ressources limitées devraient présenter des changements dans leurs indices vitaux comme une réduction de la survie des juvéniles, un ralentissement de la maturation et une réduction de la survie des adultes. Le taux de croissance de la population des phoques gris (Halichoerus grypus) de l'île de Sable, en Nouvelle-Écosse, a été suivi de 1963 à 2010 en estimant la production de petits. Récemment, le taux d'augmentation de la production de petits est passé de 12 % par année avant 1997 à 4 % par année. De 1969 à 2002, de façon périodique, plus de 7 000 phoques gris ont été marqués d'une manière unique au moment de leur sevrage. Les chercheurs de l'Institut de Bedford (D. Bowen, C. den Heyer et J. McMillan) et de l'Université Dalhousie (S. Iverson) ont recompté ces phoques gris marqués chaque année de 1983 à 2010 en utilisant de trois à cinq recensements hebdomadaires de la colonie de reproduction. Les recaptures de femelles marquées ont été utilisées dans une analyse de capture-recapture afin d'estimer la survie des juvéniles (du sevrage à l'âge de 4 ans) et les probabilités liées à l'âge (de 4 à 13 ans) des primipares. Deux groupes de cohortes (de 1985 à 1989 et de 1998 à 2002) ont été analysés séparément afin de détecter les changements temporels. Les taux estimés de survie des adultes sont toujours demeurés élevés (de 92 à 99 %), mais ils ont légèrement diminué au cours des années 1990. Par contre, les probabilités estimées de la première mise bas aux âges 4 à 13 ans n'ont pas changé, et la survie apparente des juvéniles est passée de 78 % dans les années 1980 à 35 % dans les cohortes les plus récentes (figure 4). Même si le taux de croissance de la population a ralenti de façon considérable, nos résultats indiquent que la survie des adultes est restée élevée. Par conséquent, si les restrictions de nourriture sont responsables du changement observé dans le taux de croissance, la dépendance à la densité est exprimée par la réduction marquée de la survie des juvéniles.
1.4 Reproduction des phoques annelés dans la baie d'Hudson (Steve Ferguson, Magaly Chambellant)
Les phoques annelés figurent parmi les espèces de phoques les plus petites et les plus abondantes, et ont une répartition circumpolaire septentrionale. Les animaux sexuellement matures utilisent principalement la banquise côtière stable avec suffisamment de neige pour établir des abris subnivaux essentiels à la survie des petits. Dans le cadre d'un programme de suivi basé dans les collectivités établi par le MPO de Winnipeg, des échantillons et des mesures de phoques annelés ont été recueillis par les chasseurs Inuits au cours de leur pêche de subsistance d'automne à Arviat, au Nunavut. L'âge des phoques annelés a été déterminé en étudiant les groupes de couches de croissance dans le cément de sections de canines préalablement décalcifiées, coloriées et amincies. Les ovaires et les testicules ont été congelés lorsqu'ils ont été recueillis et ils ont été examinés plus tard aux fins d'évaluation de la reproduction.
La croissance en longueur et en masse (figure 5) des phoques annelés mâles et femelles a été estimée par des courbes de croissance Gompertz. Il n'y avait aucun dimorphisme sexuel portant sur la longueur, la masse ou la condition corporelle (profondeur de la graisse) chez les phoques annelés de l'ouest de la baie d'Hudson. Toutefois, comme les femelles étaient significativement plus âgées, les mâles pourraient être un peu plus grands à un âge donné. Comparativement à d'autres emplacements dans l'Arctique, les phoques annelés de la baie d'Hudson présentaient une longueur et une masse plus petites, ce qui appuie l'hypothèse des différences de taille le long d'un gradient latitudinal.
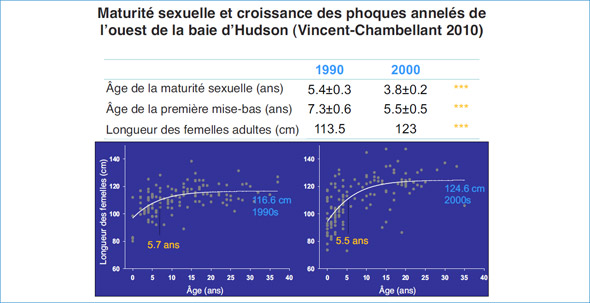
Figure 5: Résumé des changements reproductifs au cours des décennies; les années 1990 par rapport aux années 2000.
Dans la baie d'Hudson, les femelles ont atteint lamaturité sexuelle vers l'âge de six ans,mais lamaturité sexuelle morphologique a pu être détectée à partir de trois ans. Les phoques annelésmâles semblent atteindre lamaturité sexuelle vers cinq ans,mais lamasse des testicules continue d'augmenter jusqu'à l'âge de 10 et 11 ans. Les deux sexes semblent atteindre lamaturité physique avant d'atteindre leurmaturité sexuelle comportementale. Les âges de maturité sont conformes à ceux d'autres phoques annelés dans la baie d'Hudson et dans le reste de l'Arctique,même s'ils se situent dans la partie inférieure du spectre.
Le cycle reproductif des phoques annelés de la baie d'Hudson est semblable à celui des phoques d'autres emplacements de l'Arctique. Les petits naissent sur une banquise côtière ou une banquise stable dans des abris subnivaux qui nécessitent une profondeur de neige de 20 cm ou plus pour fournir une protection suffisante contre la température et les prédateurs de l'Arctique. La date de naissance nominale des petits est établie au 1er avril, mais la période de mise bas peut s'étendre sur plusieurs semaines. Dans la baie d'Hudson, les connaissances traditionnelles et les données récentes convergent vers une période de mise bas antérieure, débutant en février et dont le pic se situe à la mi-mars. Cela appuie l'hypothèse d'un gradient latitudinal des mises bas. Les petits sont sevrés avant la débâcle, après avoir été allaités durant cinq à sept semaines.
On pense que l'accouplement a lieu sous l'eau au moment du sevrage; l'activité sexuelle des mâles atteint un pic signalé de février à avril dans la baie d'Hudson. La gestation des phoques annelés prend environ 10,5 mois, ce qui comprend une période de développement suspendue au cours des premiers deux à trois mois. À la fin du printemps, les phoques annelés entreprennent leur mue annuelle et ils ont besoin d'une banquise pour s'échouer.
La condition corporelle des phoques annelés est la plus médiocre au début de l'été après avoir jeûné pendant les périodes d'accouplement et de mue. Au cours de la période des eaux libres, toutes les catégories d'âges sont mélangées et s'alimentent de façon intensive.
Lorsque la glace commence à se former à la fin de l'automne, les adultes se rassemblent près de la côte afin d'établir leurs territoires. Au cours de cette période, les jeunes sont activement exclus de ces habitats. Les phoques annelés adultes démontrent des signes de fidélité au site au cours de ces mois d'hiver et ils pourraient avoir un système d'accouplement faiblement polygyne avec défense des ressources.
Nous proposons que les dynamiques de la population des phoques annelés suivent un cycle décennal lié aux fluctuations de l'environnement et en particulier au régime de la glace de mer, par l'entremise de changements dans la productivité de l'océan et la pression des prédateurs. Dans les années 2000, les phoques plus jeunes grandissaient plus vite, atteignaient la maturité plus tôt et produisaient plus de petits ayant une meilleure survie que les phoques des années 1990. Des caractéristiques biologiques de ce genre suggèrent davantage une population croissante dans les années 2000 que dans les années 1990. Le changement des caractéristiques biologiques au cours des deux périodes d'étude appuie l'hypothèse que la dynamique de la population des phoques annelés dans l'ouest de la baie d'Hudson peut suivre les variations décennales du régime de glace de mer par l'entremise du forçage atmosphérique (p. ex., oscillation de l'Atlantique Nord). Le cycle de l'effectif des phoques annelés et du rendement reproductif est lié à la sensibilité démographique des petits à la couverture de neige et au besoin de glace stable pour leur survie. Par conséquent, un déclin à long terme de la condition physique des phoques annelés en réponse aux tendances actuelles et prévues des variables environnementales de la baie d'Hudson est probable et il renforce le besoin de données à long terme sur le mode de vie des mammifères marins qui dépendent de la glace et qui vivent à la limite sud de celle-ci.
1.5 Utilisation de la génétique pour acquérir des connaissances sur les groupes de bélugas dans la baie d'Hudson (Mike Hammill, Lianne Postma)
Partout dans leur aire de répartition, les bélugas (Delphinapterus leucas) sont connus pour visiter les estuaires et les embouchures de rivière au cours de l'été. Certains de ces estuaires sont des endroits de regroupements réguliers pour un grand nombre de baleines et ils ont traditionnellement fourni de bonnes occasions pour effectuer une chasse de subsistance. Cela s'explique par le fait que le retour des bélugas est habituellement prévisible et que de nombreuses baleines sont présentes dans un environnement clos qui facilite la chasse. En effet, l'accès fiable à ces sites de regroupement semble avoir été important dans la détermination des patrons de peuplement dans certaines régions de l'Arctique.
On a proposé plusieurs explications pour décrire ce comportement, mais aucune raison ne semble expliquer à elle seule l'existence de ces regroupements. Les estuaires semblent avoir une importance biologique et ils peuvent représenter un habitat essentiel pour ces animaux. Le retour continu des bélugas à ces emplacements et leur persistance à les occuper même s'ils font l'objet d'une perturbation répétée semblent en souligner l'importance.
Les premiers chercheurs à travailler sur ce sujet ont suggéré que ces regroupements de baleines en été étaient formés de groupes distincts d'animaux qui devaient être gérés en tant que populations distinctes ou unités de gestion distinctes. La préoccupation est que si un regroupement est trop chassé, cela peut entraîner l'abandon d'un estuaire. Ce problème peut ne pas avoir de conséquences sur la conservation de l'espèce, mais il peut entraîner une perte de la biodiversité ainsi que des ressources alimentaires pour certains villages situés dans des régions précises.
Afin de déterminer si les bélugas qui vivent dans la baie d'Hudson et dans les alentours sont réellement des groupes distincts de baleines, nous utilisons des marqueurs d'ADN afin de comparer les profils génétiques des animaux. Afin de nous aider, les chasseurs recueillent un échantillon de peau des animaux qu'ils ont chassés et ils nous l'envoient avec des renseignements sur la date et l'emplacement où l'animal a été chassé. Ensuite, nous examinons l'ADN que nous pouvons prélever de l'échantillon de peau et nous cherchons des similitudes et des différences entre les échantillons recueillis de différentes régions au cours de différentes saisons et par différents chasseurs.
Les bélugas sont observés le long de la côte est de la baie d'Hudson, dans la baie James et le long de la côte ouest de la baie d'Hudson au cours des mois d'été. Le groupe de l'est de la baie d'Hudson est le plus petit et il est formé d'environ 3 000 animaux. Les bélugas qui passent l'été à l'ouest de la baie d'Hudson se chiffrent à 57 000, et ils forment peut-être le regroupement de baleines le plus important au monde. Un total de 9 000 autres baleines se trouvent dans la baie James. Les bélugas de la baie d'Hudson migrent à la fin de l'été ou à l'automne vers le détroit d'Hudson, où ils passent l'hiver, même si certains animaux de la baie James semblent rester dans la région de la baie James tout au long de l'année. Les patrons de migration de ces baleines sont très intéressants, car il s'agit du moment où les différentes populations semblent se mélanger et cela leur donnerait l'occasion de s'accoupler. On pense que l'accouplement entre les bélugas a lieu au cours des mois de la fin de l'hiver et du début du printemps. La gestation dure de 9 à 12 mois; les baleineaux naissent le printemps suivant. Les baleineaux restent auprès de leur mère de 12 à 18 mois. Ces caractéristiques biologiques des bélugas contribuent aux types de patrons que nous pouvons constater lorsque nous examinons l'ADN des échantillons fournis par les chasseurs.

Échantillon de Muktuk
Photo: Lisa Loseto
Au cours des analyses génétiques, nous examinons deux types d'ADN : l'ADN mitochondrial et l'ADN nucléaire. L'ADN mitochondrial (ADN mt) est un type d'ADN qui est principalement transmis de la mère à son petit. L'examen des renseignements tirés de l'ADN mt est utile pour l'identification de groupes d'animaux en fonction de leur lignée maternelle. Par contre, l'ADN nucléaire (ADN n) est le type d'ADN qui est le plus fréquemment utilisé dans la science judiciaire, car il est hérité de façon égale de chacun des parents. Pour les bélugas, l'ADN n peut donc être utile pour découvrir des renseignements sur les patrons de reproduction, c.-à-d. les groupes qui s'accouplent ensemble et le type de relations que les baleines individuelles ont entre elles.
Parmi les échantillons d'ADN recueillis des bélugas des côtes ouest et est de la baie d'Hudson et des animaux passant l'hiver dans le détroit d'Hudson, il n'y a que peu de différenciation ou aucune différenciation entre les échantillons à l'aide de l'ADN nucléaire. Cela indique que les animaux qui se reproduisent entre eux le font probablement dans les régions d'hivernage où les animaux ont l'occasion d'être ensemble. Toutefois, les échantillons des animaux provenant des différentes zones d'estivage contenaient des types d'ADN mitochondrial distincts. Les différences d'ADN mt les plus importantes ont révélé que les bélugas capturés en été le long de l'arc de la côte de l'est de la baie d'Hudson (EBH) formaient un groupe génétiquement distinct d'animaux. Sur le plan génétique, ces baleines de l'EBH sont plus liées aux bélugas du fleuve Saint-Laurent et elles sont très différentes des deux principaux groupes de bélugas à proximité, c.-à-d. les baleines de l'ouest de la baie d'Hudson et du sud-est de l'île de Baffin.
À l'aide des signatures génétiques d'ADN mt, on a déterminé que les échantillons de la partie du Nord du Québec (Nunavik) du détroit d'Hudson contiennent une proportion importante (de 7 à 31 %) de bélugas du groupe de l'EBH. Cette proportion est beaucoup plus élevée que les prévisions basées sur la taille relative du groupe de baleines de l'ouest de la baie d'Hudson (57 000 animaux) comparativement au groupe de bélugas de l'est de la baie d'Hudson (3 000 animaux). Cela indique des différences dans les patrons de migration entre les deux groupes, en particulier lorsqu'ils arrivent dans les eaux du détroit d'Hudson.

Extraction d'ADN de la peau
Photo: Lianne Postma
Grâce à un travail collaboratif avec des collègues de l'Université Laval, des analyses plus approfondies de l'ADN nucléaire des échantillons de peau indiquent que les parents et les petits, ainsi que la demi-fratrie et d'autres baleines parentes, se déplacent en restant en association étroite dans l'espace et dans le temps. Des relations plus solides sont signalées entre les mères et leurs filles qu'entre les mères et leurs fils. Ces résultats suggèrent un réseau d'individus liés, en particulier parmi les femelles. Ces réseaux se déplacent entre leurs régions d'été et d'hiver. Par conséquent, les jeunes bélugas sont capables d'apprendre la route migratoire précise de leur mère et de leurs parents, car ils auront effectué de deux à quatre migrations au moment de leur sevrage. Cet apprentissage de routes migratoires précises peut avoir des implications importantes en matière de conservation et de gestion. Si les animaux ont tendance à maintenir les groupes partageant des liens de parenté et à visiter les mêmes régions de façon répétitive, la perte de ces groupes entraînera la perte des connaissances au sujet des routes migratoires. Donc, si les animaux sont éliminés d'un endroit en particulier, les chances de recolonisation de cette zone sont minimes. Cela semble être le cas dans certaines régions du Nord du Québec.
Ces aperçus fournis par la génétique sont très intéressants, et ils sont également soutenus par des renseignements tirés d'autres types d'études comme les observations locales, la télémétrie satellitaire, les analyses des isotopes stables et les comparaisons des signatures de contaminants. Ensemble, les données contribuent à établir une base de connaissances sur les bélugas qui peut être utilisée afin de prédire leur vulnérabilité à divers facteurs de stress dans leur environnement. Conséquemment, les décisions en matière de gestion peuvent être orientées afin de promouvoir la conservation de ces animaux.
2.0 Rôle des mammifères marins dans l'écosystème
2.1 Résumé du processus d'évaluation zonale sur les répercussions des phoques gris sur les populations de poissons dans l'Est du Canada (Don Bowen)
Les possibles répercussions négatives de la prédation par le phoque sur les populations de poissons d'intérêt commercial et de conservation (p. ex., morue de l'Atlantique) continuent de faire l'objet d'un débat. Un facteur de contribution à ce débat est la croissance observée des populations de phoques gris dans les eaux de l'est du Canada au cours des quatre dernières décennies et les déclins importants de plusieurs populations de poissons jusqu'au point où la pêche a été interrompue. La mortalité naturelle des poissons adultes a également été estimée exceptionnellement élevée dans ces populations de poissons effondrées et ne présentant pas de signe de rétablissement. On suppose que les phoques gris ont cinq types possibles de répercussions négatives sur les populations de proies : 1) prédation, 2) compétition pour les ressources alimentaire, 3) transmission de parasites entraînant une mortalité accrue des poissons, 4) perturbation du frai entraînant une réduction du succès reproductif et 5) d'autres répercussions indirectes sur la productivité des proies.
Pêches et Océans Canada (le MPO) a tenu un atelier constitué de deux volets afin d'examiner les répercussions des phoques sur les groupes de morues de l'Atlantique dans les eaux de l'est du Canada. Le premier atelier était axé sur la nature et la qualité des données disponibles et il a permis de déterminer les analyses de données et les études de modélisation qui pourraient être effectuées avec les données existantes pour mieux traiter du problème des répercussions des phoques sur le rétablissement des pêches commerciales (compte-rendu de Pêches et Océans Canada, 2008/021). Le deuxième atelier portait sur l'examen de ces nouvelles analyses (compte-rendu de Pêches et Océans Canada, 2009/020). Toutefois, les ateliers n'étaient pas conçus pour fournir des conseils en réponse aux questions des gestionnaires des pêches.
En octobre 2010, Pêches et Océans Canada a convoqué une réunion d'évaluation zonale de cinq jours rassemblant des scientifiques, des pêcheurs et des gestionnaires des pêches à l'échelle nationale et internationale afin de fournir des conseils scientifiques sur les questions suivantes : combien de phoques gris devraient être éliminés sur une période de cinq ans afin de diminuer considérablement la mortalité naturelle des morues du sud du golfe du Saint-Laurent et d'autres stocks de morues qui présentent une mortalité naturelle élevée? Quelles pourraient être les réponses de l'écosystème (p. ex., sur le plan de l'abondance d'autres prédateurs et de proies) à l'élimination ciblée des phoques gris, en particulier puisqu'elle pourrait avoir une incidence sur le rétablissement de la morue?
Afin de tenter de répondre à ces questions, les participants ont discuté du résultat de 32 analyses scientifiques couvrant les sujets suivants (compte-rendu de Pêches et Océans Canada, 2008/021) :
- Preuve tangible de la consommation de morue par les phoques gris;
- Preuve indirecte de la consommation de morue par les phoques gris;
- Diminution minimale de la mortalité naturelle afin de rétablir les populations de morue à des niveaux de référence;
- Changements liés à l'abondance, à la répartition et à l'écologie des phoques gris;
- Scénarios de réduction des phoques gris afin de rétablir les populations de morue;
- Exemples de contrôle de prédateurs marins importants dans d'autres régions du monde;
- Conception d'une expérience contrôlée afin de vérifier la répercussion du contrôle des phoques gris sur la mortalité de la morue du sud du golfe du Saint-Laurent.
En tentant de répondre à ces questions, la science est confrontée à des sources d'incertitude considérables. Cela s'explique par le fait qu'il est impossible de mesurer sans erreur les processus ayant une incidence sur la dynamique de la population de phoques, leur consommation de proie (y compris la morue de l'Atlantique), la dynamique de la morue et l'écosystème soutenant les phoques, la morue, d'autres prédateurs et leurs proies. L'incertitude de la mesure peut être amplifiée lorsque les scientifiques tentent d'estimer la mortalité de la morue causée par les phoques, car il y a une incertitude dans l'estimation de la population totale des phoques, l'estimation de la proportion du régime alimentaire contenant de la morue et l'estimation de la taille de la population de la morue, pour n'en nommer que quelques exemples.
Il est difficile de résumer les résultats de cinq jours de discussion en quelques paragraphes, et l'utilisation de ces analyses et discussions pour fournir des conseils sur les questions de gestion a été difficile et a donné lieu à de nombreux débats. Néanmoins, les conclusions de la réunion ont été résumées dans l'avis scientifique (2010/71). Les participants ont convenu d'un certain nombre de conclusions. Les phoques gris habitent trois écosystèmes marins de l'Atlantique au sud du chenal Laurentien, reliés les uns aux autres : le sud du golfe du Saint-Laurent (sous-secteur 4T de l'OPANO) qui gèle en hiver, ce qui entraîne la migration d'un grand nombre de populations de poissons dans des zones d'hivernage, en eaux chaudes plus profondes du Cap Breton (sous-secteur 4Vn de l'OPANO) et deux écosystèmes de la plate-forme néo-écossaise (sous-secteurs 4VsW et 4X de l'OPANO). Il existe des stocks de morue distincts dans chacun des trois écosystèmes. Tous les stocks ont indiqué des déclins d'au moins 80 % de l'abondance et ils demeurent tous bas aujourd'hui. La surpêche a réduit les stocks des sous-secteurs 4T, 4Vn, et 4VsW jusqu'à une faible abondance au début des années 1990. La surpêche a également contribué au déclin moindre du stock 4X jusqu'au milieu des années 1990. Malgré la mortalité des poissons grandement réduite, la survie des morues adultes dans le sous-secteur 4T est restée à un faible niveau au cours de cette période, et le stock a continué à diminuer. Le stock de morues du sous-secteur 4VsW a rapidement chuté à la fin des années 1980, ce qui a mené à un effondrement et ensuite à la fermeture des pêches en 1993. La biomasse du stock est restée faible pendant plus d'une décennie, mais elle a récemment démontré une augmentation et une meilleure survie. Le stock de morues du sous-secteur 4X a également connu une mortalité élevée et il a continué de diminuer après le milieu des années 1990, lorsque les pêches ont été limitées.
Au cours des dernières décennies, il y a eu des changements dramatiques dans ces écosystèmes. Les stocks et les pêches de poissons de fond du sud du Golfe ont été remplacés par des poissons démersaux de petite taille et des pêches d'invertébrés. De même, les pêches de poissons de fond de l'écosystème de l'est du plateau néo-écossais ont maintenant été remplacées par les pêches d'espèces d'invertébrés comme les crevettes et les crabes. Dans l'ouest du plateau néo-écossais, les pêches d'invertébrés ont également augmenté et sont passées à des niveaux sans précédent, mais certaines pêches de poissons de fond se poursuivent.
À des fins de gestion, la population des phoques gris est divisée en trois troupeaux selon les sites de mise bas. Le troupeau le plus important, qui est formé de 260 000 à 320 000 phoques (en fonction des hypothèses de départ des modèles de population), est situé à l'île de Sable. Le taux d'accroissement de ce troupeau est passé de 12,8 % au cours des années 1980 à environ 4 % au cours des cinq dernières années. Le troupeau du sud du golfe du Saint-Laurent est formé de 55 000 à 71 000 animaux. Le troupeau de la côte de la Nouvelle-Écosse est le plus petit des trois; il contient de 20 000 à 22 000 animaux. Les phoques de chacun de ces troupeaux sont dispersés au cours de l'année pendant qu'ils sont en quête de nourriture et ils peuvent contribuer à la colonisation de nouveaux sites de mise bas. Même si on sait peu de chose au sujet de l'abondance historique, la taille de la population actuelle est la plus grande population mesurée au cours des derniers siècles.
La détermination du régime alimentaire des phoques gris est fondée sur des méthodes indirectes, car les occasions d'observation directe de leur nourriture sont limitées. Les méthodes utilisées sont fondées sur la récupération de parties dures comme des os de l'oreille de poissons à partir du contenu de l'estomac, des intestins et des excréments ainsi que sur l'analyse de la composition chimique de gras sur les phoques et leurs proies. Chacune de ces méthodes a des points forts et des points faibles. Il est également difficile d'obtenir un échantillon représentatif du régime des phoques gris, car ils sont très dispersés et leur régime varie par sexe, par saison, par région et selon d'autres facteurs. Les analyses des sources de données susmentionnées indiquent une vaste gamme de valeurs pour le pourcentage de morue dans le régime des phoques gris; une moyenne globale de 2 à 7 % dans le sous-secteur 4VsW, et dans le sous-secteur 4T, de 1 % pour les femelles en été à 24 % pour l'unique échantillon de mâle en hiver.
Des modèles de consommation de nourriture indiquent que la consommation de morue des phoques gris au cours des dernières années se situe de 4 500 à 20 000 tonnes par année pour le sous-secteur 4T et de 3 000 à 11 000 tonnes par année pour le sous-secteur 4VsW. Ces estimations ont une variance élevée et leurs larges gammes reflètent une incertitude attribuée aux hypothèses effectuées afin de compenser les trous dans l'échantillonnage du sous-secteur 4T et le traitement des données du régime alimentaire du sous-secteur 4VsW.
L'abattage est une pratique très répandue en tant que moyen de limiter la prédation sur le bétail et la faune; il peut être très efficace pour réduire l'abondance de prédateurs. L'abattage a également été utilisé afin de réduire les espèces de phoques. Même s'il s'agit d'une pratique répandue, l'importance de la réduction des populations de phoques et la réponse des populations de proies à ces abattages ont rarement été évaluées. Les résultats d'autres programmes de contrôle de prédateurs indiquent que des conséquences inattendues dans les réseaux trophiques, qui seront difficiles à prévoir, sont néanmoins observées de façon courante. Par conséquent, une intervention dans le sud du Golfe nécessiterait d'abord une enquête approfondie des répercussions probables sur de multiples espèces d'une interaction entre les morues et les phoques dans cet écosystème; deuxièmement, il nécessiterait un programme soigneusement conçu qui comprendrait des objectifs bien énoncés et un suivi rigoureux des populations de phoques et de morues ainsi que de l'écosystème afin d'en évaluer les conséquences.

Phoque gris mâle adulte (marque M720, 24 ans) doté d'un émetteur-récepteur mobile VEMCO (en noir) et d'un émetteur satellitaire GPS d'Argos (sur la tête).
Photo: W.D. Bowen
Même s'il y a eu un accord d'envergure sur les conclusions susmentionnées, il y a eu un débat concernant les conséquences de la consommation de morue des phoques gris sur les dynamiques récentes et prévues de la morue, en particulier dans le secteur 4T. Cela s'explique en partie par l'incertitude supplémentaire liée à l'estimation de la mortalité de la morue due aux prédateurs plutôt que seulement la consommation de morue par les prédateurs. Dans le sous-secteur 4T, les phoques gris étaient parfois considérés comme étant une source importante de mortalité pour la morue de grande taille (plus de 35 cm) et d'autres poissons de fond adultes. La localisation par satellite indique que certains phoques gris, en particulier des mâles, sont en quête de nourriture là où se situent des regroupements importants de morues adultes. Des échantillons de systèmes digestifs de phoques en quête de nourriture dans des zones de regroupements d'hiver de morues contiennent une proportion relativement élevée de morues (environ 24 % dans les mâles et 10 % dans les femelles, d'après les échantillons d'intestins), et une proportion élevée (58 %) de ces morues était d'une longueur supérieure à 35 cm. D'autres personnes étaient moins convaincues que les renseignements disponibles fournissaient un appui convaincant concernant l'impact des phoques gris sur ce stock de morues.
Pour la morue du sous-secteur 4VsW, l'ampleur de la prédation des phoques gris comparativement à d'autres sources de mortalité variait grandement selon les hypothèses de plusieurs modèles de prédation. La plupart des modèles laissaient une grande partie de la mortalité sans explication et attribuaient seulement une petite partie (moins de 17 %) de la mortalité des morues à la prédation des phoques. Des données comparables ne sont pas disponibles pour la mortalité attribuable aux phoques gris chez les morues des sous-secteurs 4X et 4Vn.
Malgré la quantité considérable et la diversité des renseignements fournis pour gérer ce problème difficile, il existe toujours une grande incertitude concernant les répercussions de la prédation des phoques gris sur les dynamiques des stocks de morue dans l'Atlantique canadien.
2.2 Associations des phoques gris en mer: aperçus fournis par un nouvel enregistreur de données (Don Bowen)

Figure 6: Mouvements d'un phoque gris mâle adulte (orange) et d'un phoque gris femelle adulte (rouge) au cours de l'automne 2009; l'emplacement de rencontres en mer entre les phoques dotés d'un émetteur-récepteur mobile est indiqué par les cercles blancs.
Nous connaissons bien le comportement des pinnipèdes au cours de la saison d'accouplement. Par contre, nous connaissons peu d'éléments concernant la nature et l'étendue des interactions sociales entre les pinnipèdes lorsqu'ils sont en quête de nourriture en mer, car leur comportement ne peut être observé. Des chercheurs de l'Institut océanographique de Bedford et de l'Université Dalhousie (Damian C. Lidgard, Ian D. Jonsen et Sara J. Iverson) utilisent une nouvelle technologie acoustique afin d'examiner la nature des associations spatiales et temporelles des phoques gris lorsqu'ils se trouvent en mer. En octobre 2009, quinze phoques gris (Halichoerus grypus) adultes de l'île de Sable, au Canada, ont été dotés d'émetteurs-récepteurs mobiles VEMCO et d'émetteurs GPS satellitaires d'Argos. Les émetteurs-récepteurs mobiles transmettent l'équivalent d'une empreinte digitale acoustique, qui identifie chaque phoque individuellement et reçoit les « empreintes digitales » des autres phoques également dotés d'un émetteur-récepteur mobile. Une rencontre entre deux individus était définie comme un groupe de détections ayant eu lieu en moins de 30 minutes. Les émetteurs satellitaires d'Argos ont transmis les coordonnées GPS des phoques pour que l'emplacement des interactions puisse être déterminé. Deux états comportementaux (mouvements lents et rapides) ont été attribués aux emplacements GPS à l'aide d'un modèle espace-état. Les émetteurs ont transmis de l'information pendant environ 73 jours avec une moyenne de 95 emplacements par jour. Sur les treize phoques dotés d'un émetteur-récepteur mobile, douze appareils ont indiqué que les phoques interagissaient avec d'autres phoques en mer. Plus de 1 800 détections ont été enregistrées dans le cadre d'environ 200 rencontres (figure 6). La durée médiane d'une rencontre était d'environ 20 minutes et le nombre médian de rencontres par phoque était de 20. La répartition spatiale des états comportementaux lents et rapides a indiqué que les phoques effectuaient des mouvements lents (on pense qu'ils représentent la quête de nourriture) lorsqu'ils étaient sur des bancs au large où se trouvent les proies fréquemment consommées et qu'ils effectuaient des mouvements rapides (déplacement) entre ces emplacements. Les femelles étaient plus susceptibles de rencontrer d'autres phoques marqués lorsqu'elles effectuaient des mouvements lents, tandis que les mâles n'ont démontré aucune tendance en ce qui a trait aux deux états comportementaux. Ces données (les premières d'une étude pluriannuelle) suggèrent l'occurrence d'associations à court terme dans les zones de quête de nourriture et fournissent de nouveaux aperçus de l'écologie de la quête de nourriture chez ce carnivore marin. Le marquage des proies, comme la morue de l'Atlantique (Gadus morhua), avec des étiquettes acoustiques codées peut également constituer un moyen d'examiner les interactions entre les prédateurs et leurs proies.
2.3 Alimentation spécialisée des épaulards sur le saumon quinnat (John K.B. Ford)
Alimentation spécialisée des épaulards sur le saumon quinnat (John K.B. Ford) Les épaulards sont au sommet de la chaîne alimentaire des océans et constituent un des mammifères les plus largement distribués dans le monde. En tant qu'espèce, les épaulards peuvent être considérés comme étant des prédateurs généralistes; ils ont un régime qui comprend différents types de proies, dont les phoques, les dauphins et les baleines de grande taille, tous les types de poissons, allant des petits bancs de hareng aux requins-baleines, de nombreux types d'invertébrés, comme les calmars et les pieuvres, et même les reptiles, comme la tortue luth. De façon générale, plus de 150 espèces d'organismes marins ont été documentées comme étant des proies des épaulards.
Malgré ce régime généraliste, les études sur le terrain dans plusieurs régions du monde ont révélé que les populations locales d'épaulards peuvent avoir des régimes remarquablement spécialisés et qu'elles peuvent s'alimenter sélectivement d'un sous-ensemble très petit d'espèces de proies. Ces populations écologiquement spécialisées, ou écotypes, peuvent avoir des patrons distincts de répartition saisonnière, de structure sociale, de comportement et de vocalisations qui sont fortement influencés par leur mode de vie de prédateur spécialisé. Différents écotypes d'épaulards ont souvent des aires de répartition se chevauchant, mais ils ne se mélangent pas et ils sont donc isolés sur le plan reproductif. On pense qu'une isolation reproductive à long terme de ce genre a entraîné une divergence génétique parmi certains écotypes et on pense qu'ils représentent potentiellement des espèces distinctes.
Les études à long terme des épaulards au large de la côte ouest du Canada effectuées par des scientifiques de la Station biologique du Pacifique du MPO ont révélé l'existence de trois écotypes distincts dans la région, qu'on appelle épaulards « résidents », « migrateurs » et « océaniques ». Les épaulards résidents s'alimentent d'une variété de poissons et de certains calmars, mais leur régime est dominé par le saumon. Les épaulards migrateurs ne semblent pas s'alimenter de poissons, mais ils ciblent plutôt presque exclusivement des mammifères marins, dont les phoques, les otaries, les marsouins, les dauphins et les petites baleines. Le régime des épaulards océaniques n'est pas bien connu, mais il semble comprendre une proportion élevée de requins, ce qui peut justifier l'usure exceptionnellement grave des dents de cet écotype (la peau des requins est très abrasive). Ces trois écotypes, qui sont chacun composés de populations de quelques centaines de baleines, se partagent les eaux côtières de la Colombie-Britannique, mais on ne les voit jamais se déplacer ensemble ou se mélanger.

Troupeau d'épaulards
Photo: Brian Gisborne
Les épaulards résidents constituent l'écotype le plus connu des trois. Ces baleines se déplacent de façon saisonnière selon les patrons de migration de leur proie principale, le saumon du Pacifique. Ils ont longtemps été connus pour se regrouper dans de bonnes zones de pêche au saumon le long de la côte durant le pic estival de la migration du saumon vers les frayères en rivière, et on supposait qu'ils s'alimentaient des cinq espèces du saumon du Pacifique approximativement en proportion de la disponibilité de chaque espèce. Toutefois, des études sur le terrain portant spécifiquement sur le comportement alimentaire de ces baleines ont révélé que ce n'est pas le cas. À l'aide d'écailles et de tissus prélevés dans l'eau à l'emplacement où les saumons ont été tués, on a pu identifier les espèces et découvrir que les épaulards résidents s'alimentent de préférence sur le saumon quinnat, un des saumons les plus rares de la région. Des espèces abondantes comme le saumon rouge et le saumon rose, qui sont plus nombreux que le saumon quinnat dans une proportion de 1 000 pour un au cours de leur migration estivale, ne sont étonnamment pas importantes dans le régime des baleines.
La préférence des baleines pour le saumon quinnat peut se comprendre – il est de loin le plus grand saumon du Pacifique et il a tendance à posséder la teneur en gras et en huile la plus élevée, ce qui confère à chaque poisson une densité énergétique plus élevée que les autres saumons. De nombreux stocks de saumon quinnat passent leur vie entière dans l'habitat côtier des baleines, ce qui les rend disponibles toute l'année pour l'alimentation des baleines. Il est plus difficile d'expliquer pourquoi les baleines ne s'attaquent pas au saumon rouge et rose plus abondant. Ces espèces de saumon sont d'une taille relativement petite et elles sont seulement accessibles aux baleines pendant un court laps de temps en été lorsqu'elles traversent les eaux côtières pour se déplacer de la haute mer vers leurs rivières de frai. Au cours de cette période, le saumon quinnat migre également et il est aussi facilement accessible. Il est probable que les épaulards s'attaquent principalement au saumon quinnat tout au long de l'année, et cet accent sur le saumon quinnat se maintient même pendant l'été lorsque d'autres espèces de saumon sont plus abondantes. Les tactiques d'alimentation des baleines sont probablement bien adaptées pour la prédation efficace du saumon quinnat, et les espèces plus petites peuvent être plus difficiles à attraper; elles ne sont donc probablement pas si avantageuses.

Épaulard résident avec un saumon quinnat
Photo: Brian Gisborne
Le saumon quinnat semble être tellement important pour les épaulards résidents que l'accessibilité de cette seule espèce de proie peut être essentielle à leur survie. Après deux décennies de croissance lente, mais régulière, les deux populations distinctes d'épaulards résidents le long de la côte ouest, les résidents du Nord et du Sud ont chuté de façon importante vers la fin des années 1990. L'analyse démographique a révélé que ce déclin est principalement attribuable à une augmentation dramatique des mortalités au cours de cette période et ensuite à une réduction des taux de naissance. Même si les populations du Nord et du Sud se sont stabilisées et ont même commencé à augmenter légèrement au début des années 2000, elles ont été inscrites respectivement comme espèces en voie de disparition et menacée en vertu de la Loi sur les espèces en péril du Canada en 2003. Une analyse de l'abondance côtière du saumon quinnat au cours d'une période de 25 ans a révélé une forte corrélation entre la survie des baleines et l'abondance du saumon quinnat. L'élément le plus étonnant est que la mortalité des épaulards résidents a monté en flèche à des niveaux deux à trois fois plus élevés que prévu à la fin des années 1990, lorsque l'abondance du saumon quinnat a chuté de près de la moitié de la moyenne à long terme pendant plusieurs années consécutives.
Comme le rétablissement des épaulards résidents peut dépendre de la disponibilité suffisante du saumon quinnat, il est important d'avoir la meilleure compréhension possible des patrons saisonniers et spatiaux de la prédation de ces baleines et des effets potentiels des pêches sur l'abondance du saumon quinnat. Notre recherche est actuellement axée sur l'amélioration de notre compréhension des stocks de saumon quinnat importants pour les épaulards par l'entremise de l'identification génétique des stocks à partir de fragments de proies. Notre recherche est également axée sur l'estimation de la quantité de saumon quinnat nécessaire pour maintenir l'abondance actuelle des baleines et permettre la croissance future de la population. De plus, nous travaillons étroitement avec les scientifiques et les gestionnaires spécialistes des baleines et des saumons au sein du MPO et de la National Ocean and Atmospheric Administration (NOAA) aux États-Unis afin de déterminer si les pêches existantes ont potentiellement une incidence sur le rétablissement des épaulards résidents.
2.4 Écologie alimentaire du rorqual bleu dans l'estuaire du Saint-Laurent (Véronique Lesage, Thomas Doniol-Valcroze)
L'alimentation se situe au cœur du cycle biologique et écologique d'un animal. Les prédateurs de grande taille ne s'alimentent pas continuellement, mais plutôt au cours d'épisodes d'activité intense séparés par des périodes de recherche, de repos ou de socialisation. De plus, l'alimentation n'a pas lieu de manière aléatoire dans l'espace, car les animaux sélectionnent des régions précises ayant des caractéristiques de densité de proies, d'accessibilité et de prévisibilité qui maximisent leurs chances de satisfaire leurs besoins énergétiques. Chaque été, les rorquals bleus de la population en voie de disparition de l'Atlantique Nord viennent dans l'estuaire du Saint-Laurent s'alimenter des denses agrégations d'euphausiacés de la région. Identifier les moments et les emplacements correspondant à des recherches de nourriture fructueuses est donc très important afin d'évaluer et de suivre la qualité de l'habitat de cette zone d'alimentation.
Toutefois, dans l'environnement marin, l'alimentation a principalement lieu sous la surface et il est rare de l'observer directement. Dans le cadre de cette étude, nous avons utilisé des enregistreurs de données afin de consigner, à chaque seconde, la profondeur et la vitesse de dix rorquals bleus au cours de leurs plongées dans l'estuaire du Saint-Laurent. En détectant les changements rapides de vitesse caractéristiques de l'ouverture de la gueule, nous avons été en mesure de déterminer le moment exact, la profondeur et l'emplacement de chaque tentative d'alimentation. Grâce à ces renseignements, nous avons démontré que les rorquals bleus s'alimentent à tout moment du cycle diurne et qu'ils augmentent leur activité d'alimentation la nuit lorsque les proies sont accessibles dans des eaux peu profondes. Ceci contredit les hypothèses précédentes dans la littérature selon lesquelles les rorquals bleus ne s'alimentaient pas pendant la nuit. À l'aide de la télémesure radio, nous avons également été en mesure de décrire les habitats où les rorquals bleus concentrent leurs efforts en matière d'alimentation et comment les différents habitats étaient utilisés à différentes phases du cycle des marées (p. ex., alimentation le long des plateaux lorsque les courants de marée poussaient les euphausiacés en grande concentration contre les pentes abruptes).

Rorqual bleu
Photo: Thomas Doniol-Valcroze
De plus, nous avons démontré que les rorquals bleus du Saint-Laurent utilisaient des stratégies optimales afin d'adapter leur période de plongée et leurs efforts d'alimentation à la profondeur de leur proie. En particulier, les taux d'alimentation étaient systématiquement plus élevés lorsque les rorquals bleus effectuaient de courtes plongées d'alimentation dans des eaux peu profondes. Ces résultats suggèrent que les prédateurs qui s'alimentent en plongeant pourraient estimer la qualité de l'habitat selon l'accessibilité des proies à faibles profondeurs au lieu de sélectionner l'habitat uniquement en fonction de la densité ou de l'abondance des proies. Utilisées conjointement, ces stratégies peuvent permettre aux rorquals bleus d'optimiser une courte saison d'alimentation et maximiser l'acquisition de ressources. En effet, les taux d'alimentation ont diminué au cours de la saison d'alimentation estivale et ils étaient corrélés négativement au temps que chaque animal passe en paire avec un autre individu, ce qui suggère un compromis entre l'alimentation et la socialisation à l'approche de la saison d'accouplement. Une meilleure compréhension du comportement et de l'écologie alimentaires des baleines de grande taille peut contribuer à prévoir leurs réponses aux changements environnementaux et aux pressions anthropiques.
Ce projet a été effectué en collaboration avec Robert Michaud et Janie Giard du Groupe de recherche et d'éducation sur les mammifères marins à Tadoussac, au Québec.
2.5 Océanographie de l'aire d'alimentation des baleines à fanons : le piège océanographique (Yvan Simard)

Figure 7: Exemple du fonctionnement du « piège océanographique » de l'aire d'alimentation des baleines à fanons de l'estuaire du Saint-Laurent située dans le parc marin Saguenay-Saint-Laurent.

Une photo d'un groupe d'épaulards dans la région de l'estuaire de Churchill prise la même journée où on a observé la prédation sur des bélugas par des épaulards à 20 km à l'ouest de la baie Button, le 27 août 2011.
Photo: Pete Ewins
Qu'est-ce qui rend des zones océaniques précises particulièrement attrayantes pour l'alimentation des baleines à fanons? Plusieurs points névralgiques de l'écosystème constituent des « pièges océanographiques » pour le zooplancton, la nourriture préférée des cétacés. Un exemple du fonctionnement de ces systèmes est présenté dans la figure 7 pour l'aire d'alimentation des baleines à fanons située dans le parc marin Saguenay-Saint-Laurent.
Cette aire d'alimentation traditionnelle existait déjà lorsque les premiers chasseurs de baleines européens sont arrivés il y a 450 ans. Aujourd'hui, cette région constitue un des sites d'observation des baleines les plus importants au monde. Quels sont les processus fondamentaux responsables de la production et de la persistance à long terme de cette aire riche en nourriture, régulièrement visitée par les baleines du nord-ouest de l'Atlantique? Il s'agit de la question abordée par une équipe multidisciplinaire dans le programme de recherche écosystémique et océanographique brièvement résumé dans le présent document.
À l'aide de techniques hydroacoustiques à plusieurs fréquences, de mesures océanographiques, d'échantillonnage de plancton et de modèles de circulation en 3D à haute résolution couplés avec des modèles de comportement du krill validés sur le terrain, les résultats intégrés indiquent clairement comment les processus océaniques se combinent afin de piéger le krill dans des emplacements prévisibles, notamment à la tête de chenaux et de canyons, habitats caractérisés par une combinaison de propriétés particulières. Ce mode de fonctionnement 1, appelé « le piège océanographique », implique les éléments suivants : 1) la topographie sous-marine, 2) la forte et persistante circulation estuarienne en deux couches qui trie le krill par taille et pousse le krill adulte vers la tête du chenal, 3) le phototactisme négatif du krill afin d'éviter les prédateurs visuels, qui favorise ses migrations verticales journalières et le force à se concentrer lors de conditions de remontée d'eau ayant lieu le long des pentes à chaque cycle de marées et à se concentrer de façon intensive à la tête du chenal Laurentien, 4) l'advection à des fréquences de marée semi-diurnes et bimensuelles, tout comme à des fréquences plus basses, ce qui module le processus au complet en y imprimant sa variabilité dans les patrons d'agrégation et de dispersion du krill. Le processus d'agrégation du krill illustré par cette recherche est probablement général et applicable à d'autres aires d'alimentation de baleine avec ce mode d'agrégation de nourriture 1. Pour obtenir des détails, consulter le document Simard (2009) et les références énumérées ci-dessous.
La recherche écosystémique actuelle aborde le couplage dans le transport entre la région source du krill adulte, dans le golfe, et les agrégations à la tête du chenal dans le parc marin à l'aide d'une série d'observatoires océaniques qui suivent les courants et la biomasse du krill à divers emplacements le long de la route de transport, en continu au cours du cycle annuel. En temps utile, des indicateurs de l'état de l'écosystème des baleines à fanons pourront être élaborés en combinant les résultats des observatoires avec les modèles couplant circulation et krill.
2.6 Prédation des épaulards de l'Arctique (Steve Ferguson)
Les épaulards (Orcinus orca) ont une répartition globale, mais de nombreuses populations situées à des latitudes élevées ne sont pas bien étudiées. Des informations anecdotiques, des rapports d'observation, le savoir traditionnel Inuit et l'identification photographique indiquent que la présence des épaulards dans la baie d'Hudson augmente. Les épaulards n'étaient pas connus pour être présents dans la région avant le milieu des années 1900, mais depuis ce temps, on a constaté une augmentation exponentielle des signalements. On a observé les épaulards se nourrir de nombreuses espèces de mammifères marins dans la baie d'Hudson. On se préoccupe particulièrement de la prédation sur les baleines boréales (Balaena mysticetus) dans le bassin Foxe, le narval (Monodon monoceros) dans le nord-ouest de la baie d'Hudson et les bélugas (Delphinapterus leucas) dans le sud-ouest de la baie d'Hudson. L'impact de la prédation des épaulards sur les espèces de mammifères marins est inconnue, car il n'y a aucune étude à long terme ni observation directe du comportement de chasse de l'épaulard dans ces eaux. Nous avons effectué une enquête semi-dirigée sur le savoir écologique traditionnel afin de fournir des renseignements supplémentaires sur l'écologie alimentaire des épaulards. Les utilisateurs locaux des ressources naturelles sont des observateurs compétents de leur environnement, et les chasseurs Inuits ainsi que les aînés de la collectivité ont des connaissances approfondies sur les épaulards. En utilisant ces renseignements, en définissant les exigences énergétiques des épaulards et en considérant la démographie des populations de proies, nous pouvons commencer à évaluer les exigences de base des dynamiques prédateur-proie dans l'écosystème marin de la baie d'Hudson. Afin d'estimer l'incidence de la prédation, nous avons utilisé un modèle simple d'équilibre de masse pour les mammifères marins qui comprenait la structure d'âge, la taille de la population et les taux de prédation. Pour la région de la baie d'Hudson, les résultats du modèle ont indiqué que les épaulards ne font pas preuve d'une grande spécialisation en matière de proie; à la place, ils s'alimentent alternativement de narvals et de bélugas au début et à la fin de la saison libre de glaces. Il existe des preuves à l'appui de l'hypothèse indiquant que pendant le pic de la saison des eaux libres, la prédation des épaulards peut être axée sur les baleines boréales. La stratégie mixte d'alimentation utilisée par les épaulards comprend la spécialisation prédatrice saisonnière et elle a une importance enmatière de gestion et de conservation, car la prédation des épaulards pourrait ne pas être restreinte par une réponse fonctionnelle de régulation.
3.0 Interactions entre les mammifères marins et les humains
3.1 Les phoques communs comme indicateurs de la contamination du réseau alimentaire au Canada (Peter S. Ross, Michel Lebeuf)
Les polluants organiques persistants regroupent les produits chimiques possédant quatre propriétés clés : ils sont persistants, bioaccumulables, toxiques et assujettis à un transport de longue portée. Les polluants de cette catégorie comprennent les biphényles polychlorés (BPC), les dioxines (polychlorodibenzoparadioxines ou PCDD) et le DDT. Même si la plupart des polluants organiques persistants ont été réglementés au Canada et qu'ils sont actuellement assujettis à la Convention internationale de Stockholm de 2001, des préoccupations subsistent en ce qui a trait à la persistance de ces contaminants. L'utilisation du DDT comme insecticide a mené à une défaillance reproductive complète d'un grand nombre d'oiseaux se nourrissant de poissons en Amérique du Nord et en Europe. Les BPC ont aussi été liés à une défaillance reproductive, à l'immunotoxicité et à la perturbation d'endocrines chez les mammifères marins. Des dioxines ont également été impliquées dans la défaillance reproductive du touladi dans le lac Ontario pendant la période de 1945 à 1980.
Les polluants organiques persistants sont lipophiliques et se répartissent facilement dans les tissus adipeux des organismes vivants. La persistance métabolique de ces polluants entraîne leur bioamplification lors du passage aux niveaux trophiques supérieurs. Les espèces des niveaux trophiques élevés ont des concentrations jusqu'à un milliard de fois plus élevées que l'eau environnante. Les programmes de recherche et de surveillance écotoxicologiques fournissent un aperçu des priorités en matière de conservation pour certaines populations de mammifères marins ainsi que les risques pour la santé humaine liés à la consommation d'aliments d'origine marine. Comme ces polluants entrent dans l'environnement à partir d'une combinaison de sources ponctuelles et non ponctuelles, les études sur les mammifères marins peuvent fournir un signal intégré de contaminants dans les réseaux alimentaires et ainsi fournir des renseignements pour les règlementations, l'atténuation, la conservation et la gestion des pêches. En plus des sources locales et régionales, le transport de longue portée par l'entremise de processus atmosphériques et de migrations biologiques apporte les contaminants dans les régions côtières.

Les phoques communs s'échouent sur des rochers intertidaux dans le détroit de Georgia, en Colombie-Britannique, au Canada.
Photo: Peter S. Ross.
Les polluants organiques persistants préoccupants au Canada comprennent les BPC, les dioxines, les furanes, les pesticides organochlorés, dont le DDT ainsi que les éthers diphényliques polybromés. Des niveaux élevés de BPC ont été signalés dans les épaulards (Orcinus orca) résidents et migrateurs de la Colombie-Britannique et les bélugas (Delphinapterus leucas) de l'estuaire du fleuve Saint-Laurent. Pour ces cétacés figurant dans la Loi sur les espèces en péril (LEP), les polluants organiques persistants ont été identifiés à des niveaux préoccupant à l'égard de la conservation des espèces dans le contexte des programmes de rétablissement. Même si les polluants organiques persistants ont diminué chez les mammifères marins du Canada depuis la mise en œuvre des règlements au cours des années 1970, l'émergence de contaminants incluant les éthers diphényliques polybromés démontre une augmentation constante chez les mammifères marins du début des années 1980 jusqu'au début des contrôles partiels en 2004.
Les phoques communs (Phoca vitulina) sont de petits pinnipèdes qui sont largement répartis dans l'ensemble de l'hémisphère Nord. Ils se situent sur les côtes du Pacifique et de l'Atlantique du Canada ainsi que dans l'Arctique et l'estuaire du fleuve Saint-Laurent (figure 8). Ils sont considérés comme étant omnivores, mais ils préfèrent les petites proies riches en lipides, dont le merlu (Merluccius sp.), le hareng (Clupea sp.) et le poulamon (Microgadus sp.). Le phoque commun est devenu un indicateur utile de la contamination du réseau alimentaire marin en raison de son abondance, de sa répartition, de son niveau trophique élevé, de sa nature non migratoire et de sa relative facilité de manipulation.

Figure 8: Les phoques communs (Phoca vitulina) sont des pinnipèdes qui sont largement répartis le long des côtes du Pacifique, de l'Atlantique et de l'Arctique du Canada (zones ombrées).
Afin de caractériser les niveaux et les risques de contamination de cette espèce, des jeunes phoques communs ont été capturés vivants sur des sites d'échouerie à l'aide de filets de capture et de petites embarcations. Des prélèvements de gras, de peau et de sang ont été recueillis dans des conditions aseptiques. Les échantillons ont été conservés pour l'analyse des BPC (n = 160) et des éthers diphényliques polybromés (n = 44) au Laboratoire d'expertise pour l'analyse chimique aquatique (LEACA) à Sidney, en C.-B.
Les résultats ont révélé des concentrations modérées de BPC et d'éthers diphényliques polybromés se trouvant dans les phoques des quatre régions de l'étude (figure 9). Malgré les règlementations, les BPC ont des concentrations plus élevées que les éthers diphényliques polybromés. Les niveaux de BPC étaient plus élevés chez les phoques de l'estuaire du fleuve Saint-Laurent, reflétant ainsi la proximité des sources industrielles de cette région. D'un autre côté, les concentrations d'éthers diphényliques polybromés étaient plus élevées chez les phoques du détroit de Georgia, en Colombie-Britannique, reflétant probablement la proximité des municipalités majeures connues pour déverser des éthers diphényliques polybromés par l'entremise d'effluents d'eaux usées (p. ex., Vancouver). Malgré les niveaux relativement faibles de BPC et d'éthers diphényliques polybromés chez les phoques de Terre-Neuve-et-Labrador provenant de contributions locales, les résultats démontrent clairement la répartition omniprésente de ces polluants organiques persistants dans les réseaux alimentaires côtiers.

Figure 9: Les phoques communs de quatre régions du Canada révèlent la contamination des réseaux alimentaires côtiers par les BPC et les éthers diphényliques polybromés. Les BPC continuent d'être présents chez les espèces ayant un niveau trophique élevé malgré les règlementations établies au cours des années 1970. Les éthers diphényliques polybromés font actuellement l'objet d'une surveillance minutieuse réglementaire à la suite d'augmentations rapides dans les organismes vivants.
Les concentrations de BPC et d'éthers diphényliques polybromés dans l'environnement marin reflètent des historiques différents d'utilisations, d'émissions et de règlementations, et leurs conséquences sur les réseaux alimentaires adjacents. Les phoques communs fournissent une mesure intégrée de la contamination du réseau alimentaire côtier, ce qui permet d'obtenir un aperçu des contaminants préoccupants dans l'environnement côtier. Les résultats fournissent de l'information pour la gestion de l'écosystème et contribuent à établir les priorités des règlementations, des objectifs de conservation ainsi que des autres stratégies de gestion.
3.2 Relevés des baleines boréales et marquage par satellite : mer de Beaufort, 2007-2010 (Lois Harwood)
Les baleines boréales appartenant à la population de la mer de Béring (également appelée la population B-T-B, de la région de Béring-Tchoukotka-Beaufort) passent l'hiver dans la mer de Béring, pour ensuite retourner chaque année à leur aire d'estivage dans la mer de Beaufort canadienne et le golfe d'Amundsen. Certains sites de la mer de Beaufort utilisés par les baleines boréales en quête de nourriture comprennent des régions qui ont été assujetties à des activités d'exploration d'hydrocarbures dans les années 1980 et, plus récemment, où on a effectué des relevés sismiques 2D et 3D en 2001-2002 et en 2006-2010. Même s'il n'y avait aucune activité sismique extracôtière dans la mer de Beaufort en 2011, au moins deux programmes sont prévus pour 2012. Il faut également noter que la route de migration d'automne de la baleine boréale croise des emplacements d'intérêt pour l'industrie des hydrocarbures dans la mer de Beaufort d'Alaska et la mer de Tchoukotka centrale, qui comprend la concession 193 actuellement active.
Pendant le mois de juillet, les baleines boréales sont largement dispersées dans la partie extracôtière de la mer de Beaufort canadienne, seules ou en petits groupes (deux ou trois animaux sont aperçus à la surface). Au début et au milieu du mois d'août, les baleines se rassemblent pour s'alimenter dans des zones précises et récurrentes dans l'aire de répartition d'été. Les regroupements durant la période d'alimentation ont lieu lorsque les conditions océanographiques favorisent la concentration de leur proie principale : le zooplancton (crustacés). Elles doivent chercher des zones où les proies sont adéquatement concentrées afin de satisfaire leurs besoins énergétiques. Les caractéristiques océanographiques qui mènent à la remontée des eaux riches en nutriments sont particulièrement importantes dans la détermination de la répartition du zooplancton. Dans le sud-est de la mer de Beaufort, l'échantillonnage du plancton à proximité des baleines boréales qui s'alimentent a révélé la présence principale de copépodes (de 76 à 92 %) (Limnocalanus macrurus, Calanus hyperboreus, Calanus glacialis) ainsi que des amphipodes de la famille des gammaridés et des hypéridés, de krill, de mysidacés et d'isopodes en tant que proies majeures.
Des relevés aériens de baleines boréales et des études de marquage ont été effectués afin de mettre à jour nos connaissances sur la répartition, l'utilisation de l'habitat et sur le comportement de cette baleine dans le sud-est de la mer de Beaufort. L'étude du comportement a été effectuée en collaboration avec des scientifiques et des chasseurs de l'Alaska. Les contributions apportés par ces travaux étaient de signaler l'emplacement de baleines boréales en temps réel et de signaler les observations venant des relevés aériens aux autres scientifiques qui travaillaient sur un projet d'échantillonnage océanographique complémentaire. Ces travaux comprenaient aussi une stratégie d'atténuation des effets potentiels du bruit sismique sur les baleines boréales dans le sud-est de la mer de Beaufort.
Des relevés aériens systématiques ont été menés au-dessus de la mer de Beaufort canadienne à la fin du mois d'août en 2007, 2008 et 2009 afin d'examiner la répartition des baleines boréales (Balaena mysticetus). Un total de 24-26 transects (N-S) ont été effectués en avion lors de chaque relevé, tous dans des conditions favorables, le long de lignes espacées de 15 minutes de longitude. Un total de 334 baleines boréales (244 signalements), dont 10 baleineaux, ont été observées sur les relevés, le plus souvent des individus solitaires (76,6 %) ou en groupes de deux (14,3 %). La zone d'étude était ensuite divisée en cellules de 20 km par 20 km, équivalant à l'espacement des transects. L'extrapolation du nombre de baleines visibles sur les transects aux zones de chaque cellule n'ayant pas été couvertes par les relevés a donné environ 1 320 (95 % EC de 1 036 à 1 603) baleines boréales visibles à la surface au cours du relevé de 2007. Après l'application d'une correction approximative pour les baleines submergées, on estime que 5 280 baleines boréales, c'est-à-dire au moins la moitié des estimations de la taille actuelle du stock, étaient présentes dans la zone d'étude au moment du survol.

Marquage d'une baleine boréale
Photo: DFO
Les baleines boréales étaient regroupées à neuf emplacements géographiques à l'intérieur de la zone d'étude dans le cadre des relevés de 2007 à 2009. Les neuf emplacements n'ont cependant pas tous été fréquentés chaque année, et jamais plus de six d'entre eux n'étaient fréquentés au cours d'une même année. Les eaux peu profondes au large de la péninsule Tuktoyaktuk étaient attrayantes pour les baleines boréales lors des relevés de 2007 à 2009 représentant ainsi 47,3 % des baleines observées, ce qui équivaut à 66,5 % de toutes les observations. Les huit autres emplacements où les baleines boréales étaient regroupées comprenaient une proportion variant de 1,5 % à 6,3 % des observations totales des relevés. L'importance totale (28,1 %) de ces emplacements pour les baleines boréales n'était pas égale à celle de la péninsule Tuktoyaktuk quant au nombre de baleines aperçues. La tendance des baleines boréales à se regrouper et les connaissances actuelles des zones de regroupement fréquentées au cours d'une saison donnée, fournissent un cadre de travail pour l'établissement de mesures d'atténuation liées aux relevés sismiques de la mer de Beaufort.
En 2005, l'Alaska Department of Fish and Game a entamé un projet de recherche collaborative afin d'étudier les mouvements et l'utilisation de l'habitat du stock de baleines boréales (Balaena mysticetus) de l'ouest de l'Arctique. En 2007-2010, des scientifiques du MPO et des chasseurs de baleines canadiens ont collaboré sur ce projet afin de déployer des émetteurs satellites sur des baleines boréales au Canada. Au total, 59 émetteurs satellites ont été installés en Alaska et au Canada entre 2006 et 2010, dont 21 émetteurs dans les eaux canadiennes. Ce marquage couvrant plusieurs années consécutives nous a permis d'examiner la variabilité des mouvements, des zones d'hivernage et la variabilité de la période de migration. Nous avons déterminé plusieurs zones d'utilisation intensive dans l'aire de distribution des baleines boréales, en plus d'avoir documenté des interactions avec l'activité sismique extracôtière. Des analyses plus détaillées sont en cours. Elles comprendront les mouvements des animaux par rapport aux activités industrielles et les facteurs océanographiques ayant une incidence sur les mouvements et l'alimentation des baleines boréales.
4.0 Relations avec les conseils de cogestion
4.1 Recherche sur les mammifères marins dans le Nord : Travailler avec les chasseurs et les collectivités (Becky Sjare, Lois Harwood, Steve Ferguson, Véronique Lesage)
Contexte de la cogestion
Les approches de cogestion constituent le fondement de la conservation des ressources de mammifères marins et de poissons dans la plupart des régions du Nord du Canada. En termes généraux, la cogestion est un processus législatif qui fait en sorte que les chasseurs et pêcheurs locaux, les membres de la collectivité, les organismes gouvernementaux et les divers conseils publics partagent les responsabilités en matière de gestion et de conservation des ressources de mammifères marins et de poissons. Certaines caractéristiques clés de cette cogestion sont : l'élaboration partagée de projets de recherche, la mise en œuvre et la participation de la collectivité, la partage des pouvoirs décisionnels, l'utilisation du savoir traditionnel en parallèle aux résultats des recherches scientifiques. L'intégration de toutes ces caractéristiques favorise les programmes de recherche pour qu'ils soient efficaces et significatifs pour l'ensemble des participants tout en traitant des besoins en matière de gestion et de conservation à long terme pour l'espèce en question.
La plupart des organismes de cogestion des régions du Nord ont été créés dans le cadre des accords sur les revendications territoriales d'Inuvialuit, du Nunavik, du Nunavut et du Nunatsiavut. À l'heure actuelle, il y a plusieurs conseils et comités en fonction au Canada. Les principaux organismes pertinents pour la recherche des mammifères marins par le MPO sont les suivants : 1) le Comité mixte de gestion de la pêche (CMGP) actif dans le delta du Mackenzie et la mer de Beaufort depuis 1986; 2) le Conseil de gestion des ressources fauniques du Nunavut (CGRFN) établi pour la région de l'est de l'Arctique en 1993; 3) le Torngate Joint Fisheries Board (TJFB) créé pour le nord du Labrador en 2005; 4) le Conseil de gestion des ressources fauniques de la région marine du Nunavik (CGRFRMN) établi en 2009 pour le nord du Québec.
Les organismes de cogestion sont les principaux instruments de gestion des mammifères marins (et des pêches) touchant les diverses zones des accords sur les revendications territoriales. Ils fournissent également une orientation et un cadre de travail à l'élaboration de partenariats ou de collaborations en matière de recherche avec le MPO, ainsi qu'un financement partiel ou total des travaux. Bien que la structure de gouvernance et les pouvoirs décisionnels de ces organismes varient, ils sont généralement tous composés de personnes nommées par les gouvernements et organisations autochtones, les collectivités, le MPO et d'autres ministères/organismes fédéraux. Dans certains cas, des gouvernements provinciaux ou territoriaux participent aux nominations.
Trouver un terrain d'entente
La nature et la portée des projets de recherche des mammifères marins entrepris par les organismes de cogestion et le MPO dans les régions du Nord varient grandement. Certains partenariats et certaines collaborations remontent au milieu des années 1980, comme le CMGP dans l'ouest de l'Arctique. Malgré ces différences, lors des dernières années certains thèmes de recherche sont communs aux zones visées par les différents accords, tels que : 1) l'élaboration de programmes d'échantillonnage pour les espèces concernées par des chasses de subsistance importantes d'un point de vue culturel et écologique; 2) l'amélioration de notre compréhension collective des mouvements saisonniers des mammifères marins et l'identification des habitats essentiels; 3) la compréhension et la surveillance des effets des changements climatiques sur les mammifères marins et leurs habitats; 4) l'évaluation des répercussions potentielles des activités de l'industrie sur des espèces clés; 5) l'élaboration d'une capacité de recherche, d'éducation fondée sur la science et d'une expertise en matière de prise de décision au niveau de la collectivité.
Le CMGP de l'ouest de l'Arctique était le premier organisme de cogestion à adopter un programme d'échantillonnage biologique basé sur la participation des chasseurs afin de résoudre des problèmes touchant la collectivité et de répondre au besoin d'études des mammifères marins à long terme. Le comité et les chercheurs du MPO ont reconnu que la pêche ou la chasse de subsistance des mammifères marins par les Inuvialuits pouvait fournir des données biologiques à long terme beaucoup moins coûteuses. De plus, ces données auraient été logistiquement difficiles à recueillir. Des projets collectifs quasi permanents découlant des accords dans la région des Inuvialuits comprennent la surveillance de la chasse de subsistance des phoques annelés (afin d'examiner la condition corporelle, la reproduction ; la présence de contaminants et de maladies) et de la chasse des bélugas (suivi de la taille et du sexe des baleines récoltées; suivi de la période de chasse; récolte de statistiques). La participation annuelle des collectivités du Nunavut est devenue importante pour le programme de suivi dans la région environnante de la baie d'Hudson depuis 2003. Les chasseurs du Nunavumiut recueillent principalement des échantillons de tissus de phoque (annelé, barbu, du Groenland, commun), du béluga et du narval. Ils fournissent également des échantillons des principales espèces consommées par les mammifères marins. Depuis 2008, le Torngate Joint Fisheries Board appuie le programme de collecte d'échantillons du Labrador sur les phoques annelés par les collectivités de Nain, de Makkovik, de Hopedale et de Rigolet. Un programme similaire est susceptible d'être entrepris au Nunavik lorsque le CGRFRMN nouvellement créé sera complètement établi. Ces programmes font participer l'ensemble des collectivités au suivi, à la collecte d'échantillons, à la planification, à la communication et à l'interprétation des données.

Un chasseur d'Arviat, dans la baie d'Hudson, donne une présentation aux enfants de l'école élémentaire locale. Il leur explique comment il coordonne la collecte d'échantillons de phoques annelés par les chasseurs locaux de la région et comment il examine certains échantillons (on voit un fœtus de phoque annelé).
Photo: DFO
La compréhension des mouvements saisonniers des espèces de mammifères marins est nécessaire afin de bien évaluer l'abondance de la population, de déterminer les stocks ou les unités de gestion, de déterminer les habitats essentiels et enfin pour élaborer des stratégies de gestion et de conservation à long terme. Les organismes de cogestion de toutes les régions visées par les accords ont reconnu cette nécessité. Ils collaborent avec le MPO sur divers programmes de marquage (émetteur satellites) de mammifères marins (p. ex., phoques annelés, bélugas, narvals, baleines boréales). Les contributions et participations de nombreux chasseurs aux programmes de captures d'animaux vivants notamment pour la sélection des emplacements, des périodes, des techniques de capture, de manipulation d'animaux, ainsi que leurs connaissances du comportement des animaux, ont grandement contribué à la réussite des projets. Certains résultats des projets ont surpris les chasseurs et les chercheurs du MPO et impliquent de nouvelles interprétations des comportements de migration et de plongée des animaux, en plus de redéfinir l'utilisation de l'habitat par certaines espèces. Dans plusieurs communautés on retrouve des équipes expérimentées pour le marquage de mammifères marins : à Ulukhaktok, à Tuktoyaktuk et à Aklavik dans l'ouest de l'Arctique, à Sanikiluaq, à Igloolik et à Kuujjuaraapik dans la région de la baie d'Hudson, ainsi qu'à Nain dans la région du lac Melville au Labrador.
Les impacts du changement climatique sur les mammifères marins, constituent une préoccupation majeure pour les organismes de cogestion, les collectivités et le Ministère, en particulier pour les espèces qui ont une importance culturelle, nutritionnelle et commerciale. Les données recueillies sur de longues périodes par les divers programmes de surveillance découlant des accords sont actuellement analysées. Ces analyses tiennent compte des changements climatiques dans l'environnement marin (p. ex., des changements dans la couverture de glace et d'accumulation de neige). Elles incluent aussi les données scientifiques historiques et le savoir traditionnel touchant les changements liés à la distribution des animaux et à leur abondance. Ces approches de recherche fournissent des aperçus des changements propres à une espèce. Cependant elles ne définissent pas bien pour le moment comment les dynamiques de l'écosystème sont touchées. Dans la région de la baie d'Hudson du Nunavut, des recherches visant à décrire le réseau alimentaire complet de la baie sont présentement en cours. Elle sera utilisée dans le but d'établir un modèle d'interactions trophiques des mammifères marins incluant les nutriments et le phytoplancton. Ce modèle aidera à prévoir l'incidence des perturbations du système sur les mammifères marins (p. ex., les effets des eaux plus chaudes sur les poissons proies). La compensation des répercussions des changements climatiques de l'Arctique sur les mammifères marins par la protection des habitats essentiels saisonniers et par l'identification des populations les plus vulnérables, sont des mesures pertinentes pour la conservation de ces espèces. De plus, ces renseignements peuvent être utilisés par les habitants du Nord afin de s'adapter pour préserver leurs activités culturelles et économiques découlant de la présence des mammifères marins proche de leurs communautés.

Une équipe de marquage se préparant à poser un émetteur satellite sur un jeune phoque annelé capturé dans la région du lac Melville, au Labrador.
Photo: DFO

Des membres de l'équipe de marquage de Paulatuk dans l'ouest de l'Arctique relâchent un phoque annelé équipé d'un émetteur satellite.
Photo: DFO

Un chasseur de la région de la baie d'Hudson visite le laboratoire des mammifères marins de l'Institut des eaux douces du MPO.
Photo: DFO
Les répercussions du développement industriel de grande envergure dans les régions du Nord constituent une préoccupation importante pour les tous partenaires de cogestion. Le CMGP et les chercheurs du MPO dans l'ouest de l'Arctique ont mené un certain nombre d'études conjointes, examinant les effets possibles de la production pétrolière et gazière dans la mer de Beaufort. Des projets récents ont été réalisés sur les impacts des activités de forage sur la répartition et les mouvements des phoques annelés (marquage); la répartition, les mouvements et le comportement des baleines boréales durant l'alimentation estivale dans le sud-est de la mer de Beaufort (relevés aériens et marquage) et la répartition des bélugas dans l'estuaire du fleuve Mackenzie et dans les régions extracôtières adjacentes (relevés aériens, surveillance acoustique). D'éventuels projets d'exploration pétrolière et gazière dans la baie de Baffin et à Lancaster Sound, de nouveaux projets miniers de grande envergure incluant le transport par voie maritime durant l'hiver, des projets hydroélectriques dans la région de la baie d'Hudson, ainsi que des développements hydroélectriques, pétroliers et gaziers proposés le long de la côte du Labrador, peuvent tous avoir des répercussions sur les communautés et leurs activités. Ainsi, elles constitueront une préoccupation future pour les communautés et le Ministère.
Les occasions pour les chasseurs et les autres membres de la collectivité d'acquérir de nouvelles compétences techniques et de nouvelles aptitudes de leadership, de participer à toutes les étapes d'un programme de recherche et d'offrir leur savoir écologique local de façon importante et pertinente, se manifestent clairement dans la large gamme de programmes brièvement décrits dans le présent document. De plus, comme mentionné précédemment, la réussite de nombreux programmes résulte directement de la participation des chasseurs ou de la collectivité. Dans toutes les régions visées par les accords, on a observé une mobilisation, parfois très novatrice, afin d'améliorer la communication et le transfert d'informations en tous genres, favorisant ainsi de nouvelles occasions d'échanger des connaissances entre les organismes de cogestion, les collectivités et le Ministère. Par exemple, des cartes et des animations des mouvements et des phoques (parfois en temps réel) provenant des projets de télémétrie satellitaire près de Sanikiluaq, de la baie d'Hudson, du golfe d'Amundsen, des mers de Beaufort/Tchoukotka et du lac Melville, au Labrador, peuvent être consultés sur Internet. L'affichage de ces résultats sur le Web rend l'information plus rapidement accessible à la communauté scientifique, aux médias, aux collectivités du Nord, au public, aux étudiants et aux professeurs. À l'avenir, le Ministère et les organismes de cogestion continueront à travailler étroitement ensemble afin d'assurer l'exploitation durable et la conservation des ressources de mammifères marins dans les régions du Nord du Canada.
5.0 Espèces en péril
5.1 Pratiques d'immersion en mer et habitat essentiel de l'épaulard de la Colombie-Britannique (Peter S. Ross)
Les populations d'épaulards résidents du nord et du sud de la Colombie-Britannique sont inscrites respectivement comme espèces menacées et en voie de disparition en vertu de la Loi sur les espèces en péril (LEP). Cette loi vise à empêcher les espèces en péril d'être tuées ou blessées (article 32) tout en protégeant leur habitat essentiel (article 58). On a découvert que ces épaulards résidents étaient contaminés par des niveaux très élevés de biphényles polychlorés (BPC). Ces produits chimiques hydrophobes sont intégrés dans la partie lipidique du réseau alimentaire, où ils font l'objet d'une bioamplification. Ils peuvent aussi s'attacher aux particules et aux sédiments. Des concentrations élevées de BPC découvertes dans les ports et les zones industrielles peuvent être la source de contamination des réseaux alimentaires adjacents.
Environnement Canada réglemente les activités d'immersion en mer en vertu de la Loi canadienne sur la protection de l'environnement (1999) (LCPE de 1999; Règlement de l'immersion en mer, Règlement sur les demandes de permis pour l'immersion en mer). Le MPO est chargé de l'émission de permis liés à la LEP pour les activités pouvant blesser une espèce en péril ou détruire son habitat essentiel. Un modèle du réseau alimentaire été élaboré afin d'évaluer la présence et les transferts de BPC. Il examine aussi des scénarios d'immersion et permet d'améliorer les pratiques de gestion des risques liées aux épaulards et à leur habitat essentiel.

Épaulards au large de la côte de la Colombie-Britannique
Photo: B. Gisborn
L'immersion de matériel ayant des concentrations de BPC (sédiment) à des niveaux ambiants inférieurs au site, ne devrait pas contribuer à augmenter les BPC chez les épaulards sur ce même site. Pour une immersion de matériel contenant des niveaux de BPC ambiants supérieurs à ceux retrouvés à un site, une stratégie de rechange serait l'immersion dans un autre site ayant un taux de sédimentation naturelle élevé, à condition que ce nouveau site soit réputé comme un lieu de dépôt stable et que les matières de dragage ne soient pas plus dispersées qu'elles ne l'étaient à l'origine. Ce modèle indique qu'un niveau cible de sédiment (BPC) avec un poids sec de 0,012 à 0,200 μg kg-1 protégerait mieux les épaulards, espèce ayant une longue espérance de vie et un niveau trophique élevé. Toutefois, les niveaux de sédiment de BPC acceptables sont actuellement dépassés dans de nombreuses régions côtières de la C.-B., ce qui illustre l'héritage laissé par les BPC utilisés il y a plusieurs décennies.
Disposal of materials with less than ambient sediment PCB concentrations at a receiving site is not expected to increase PCB delivery to killer whales. Disposal of materials containing greater than ambient PCB levels would benefit from an alternative strategy, including disposal at a site with high natural sedimentation rate, as long as this new site is deemed to be depositional and dredge materials are not further dispersed. This modeling effort suggests that a target sediment PCB range of 0.012 to 0.200 μg•kg-1 dry weight would optimally protect long-lived, high trophic level killer whales. However, these protective sediment PCB values are currently exceeded in many parts of coastal BC, underscoring the legacy left by the PCBs used decades ago.
En comprenant la présence, le transfert, l'origine et les taux de BPC retrouvés dans les sédiments des eaux côtières, et en connaissant les types de substrat dans les sites de dragage et d'immersion, les décisions fondées sur la gestion des risques concernant les conséquences des activités d'immersion à l'intérieur de l'habitat essentiel des épaulards seront plus éclairées.
6.0 Publications de 2009-2011
- Addison, R.F., D.C.G. Muir, M.G. Ikonomou, L. Harwood and T.G. Smith. 2009. Hexachlorocyclohexanes (HCH) in ringed seal (Phoca hispida) from Ulukhaktok (Holman), T: trends from 1978 to 2006. Science of the Total Environment 407 (2009), pp. 5139-5146. 10.1016/j.scitotenv
- Alava, J.J., Ikonomou, M.G., Ross, P.S., Costa, D., Salazar, S., Aurioles-Gamboa, D., and Gobas, F.A.P.C. 2009. Polychlorinated biphenyls and polybrominated diphenyl ethers in Galapagos sea lions (Zalophus wollebaeki). Environ.Toxicol.Chem. 28: 2271-2282.
- Alava, J.J., Ross, P.S., Ikonomou, M.G., Cruz, M., Jimenez-Uzcategui, G., Dubetz, C., Salazar, S., Costa, A.N., Villegas-Amtmann, S., Howorth, P., and Gobas, F.A.P.C. 2011a. DDT in endangered Galapagos sea lions (Zalophus wollebaeki). Mar.Pollut.Bull. 62: 660-671.
- Alava, J.J., Salazar, S., Cruz, M., Jimenez-Uzcategui, G., Villegas-Amtmann, S., Paez-Rosas, D., Costa, A.N., Ross, P.S., Ikonomou, M.G., and Gobas, F. 2011b. DDT strikes back: Galapagos sea lions face increasing health risks. Ambio 40: 425-430.
- Andersen, J. M., Y. F. Wiersma, G. Stenson, M. O. Hammill and A. Rosing-Asvid. 2009. Movement Patterns of Hooded Seals (Cystophora cristata) in the Northwest Atlantic Ocean during the Post-Moult and Pre-Breed Seasons. J. Northw. Atl. Fish. Sci., Vol. 42: 1–11.
- Anderwald, P., Daníelsdóttir, A.K., Haug, T., Larsen, F., Lesage, V., Reid, R., Víkingsson, G.A., Hoelzel, A.R. 2011. Possible cryptic stock structure for minke whales in the North Atlantic; implications for conservation and management. Biological Conservation 144: 2479-2489.
- Applebee, A., A.R. Thompson, L.N. Measures and M.E. Olson. 2010. Giardia and Cryptosporidium in harp and hooded seals from the Gulf of St. Lawrence, Canada. Veterinary Parasitology 173:19-23.
- Archambault, P., Snelgrove, P., Fisher, J.A.D., Gagnon, J.-M., Garbary, D. J., Harvey, M., Kenchington, E., Lesage, V., Levesque, M., Lovejoy, C., Mackas, D. McKindsey, C.W., Nelson, J., Pépin, P., Piché, L., Poulin, M. 2010. From Sea to Sea: Canada’s Three Oceans of Biodiversity. PLoS ONE 5(8):e12182. doi:10.1371/journal.pone.0012182.
- Asselin, N.C. and Richard, P.R. 2011. Results of narwhal (Monodon monoceros) aerial surveys in Admiralty Inlet, August 2010. CSAS Research Document 2011/056: 30 p.
- Asselin, N.C., Barber, D.G. Stirling, I., Ferguson, F.H. and Richard, P.R. 2011. Beluga (Delphinapterus leucas) habitat selection in the eastern Beaufort Sea in spring, 1975-1979. Polar Biology DOI 10.1007/s00300-011-0990-5: 16 p.
- Bahoura, M, and Simard, Y. 2009. Blue whale calls classification using short-time Fourier and wavelet packet transforms and artificial neural network. Digital Signal Processing 20: 1256-1263. doi:10.1016/j.dsp.2009.10.024.
- Bailleul, F., Lesage, V. and M. O. Hammill. 2010. Spherical First-Passage Time: a tool to investigate area-restricted search in three-dimensional movements. Ecological Modelling 221: 1665-1673.
- Bajzak, C., M.O.Hammill, G.B.Stenson and S.Prinsenberg. 2011. Drifting away: implications of changes in ice conditions for a pack ice-breeding phocid, the harp seal (Pagophilus groenlandicus).Can. J.Zool.89(11):1050-1062.
- Bajzak, C.E., S.D. Côté, M.O. Hammill, G. Stenson. 2009 . Intersexual differences in the postbreeding foraging behaviour of the Northwest Atlantic hooded seal. Marine Ecology Progres Series 385:285-294.
- Barlow, J., Calambokidis, J., Falcone, E.A., Baker, C.S., Burdin, A.M., Clapham, P.J., Ford, J.K.B., Gabriele, C.M., LeDuc, R., Matilla, D.K., Quinn, T.J. II, Rojas-Bracho, L., Straley, J.M., Taylor, B.L., Urbán R., J., Wade, P., Weller, D., Witteveen, B.H., Yamaguchi, M. 2011. Humpback whale abundance in the North Pacific estimated by photographic capture-recapture with bias correction from simulation studies.Marine Mammal Science 27(4):793-818.
- Bell, R.K., and Harwood, L.A. In press. Harvest-based Monitoring in the Inuvialuit Settlement Region: Steps for Success. Arctic.
- Benjamins, S., Kulka, D.W., and Lawson, J.W. 2010. Recent incidental catch of sharks in gillnet fisheries of Newfoundland and Labrador, Canada. Endangered Species Research 11: 133-146.
- Bennett, E.R., Ross, P.S., Huff, D., Alaee, M., and Letcher, R.J. 2009. Chlorinated and brominated organic contaminants and metabolites in the plasma and diet of a captive killer whale (Orcinus orca). Mar.Pollut.Bull. 58: 1078-1095.
- Benoit, D., Simard, Y., Gagné, J.A., Geoffroy, M. and Fortier, L. 2010. From polar night to midnight sun: photoperiod, seal predation, and the diel vertical migrations of polar cod (Boreogadus saida) under landfast ice in the Arctic Ocean. Polar Biology 33(11): 1505-1520. DOI 10.1007/s00300-010-0840-x
- Benoît, H.P., Swain, D.P., Bowen, W.D., Breed, G.A., Hammill, M.O., Harvey, V. 2011. Evaluating the potential for grey seal predation to explain elevated natural mortality in three fish species in the southern Gulf of St. Lawrence. Marine Ecology Progress Series 442:149-167.
- Benoît, H. P., Swain, D. P., and Hammill, M.O. 2011. Seasonal patterns in the spatial overlap of southern Gulf of St. Lawrence cod and grey seals, with a discussion of sources of error and possible bias. DFO Can. Sci. Adv. Sec. Res. Doc. 2011/018.
- Benoit, H.P., M.O. Hammill and D.P. Swain. 2011. Estimated consumption of southern gulf of St. Lawrence cod by grey seals:bias, uncertainty and two proposed approaches. CSAS Res. Doc. 2011/041
- Bowen, W. D. 2011. Historical Grey Seal Abundance and Changes in the Abundance of Grey Seal Predators in the Northwest Atlantic. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/026.
- Bowen, W. D. and Iverson, S. J. 2011. Sources of Bias and Uncertainty in Seal Diet Composition: Hard Part and Fatty Acid Analysis. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/025.
- Bowen, W. D. and Lidgard, D. 2011. Vertebrate Predator Control: Effects on Prey Populations in Terrestrial and Aquatic Ecosystems. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/028.
- Bowen, W. D. and Northridge, S. 2010. Morphometrics, age estimation, and growth, p. 98-118. In Boyd, I. L., Bowen, W. D., and Iverson, S. J. (eds.) Marine Mammal Ecology and Conservation: A handbook of techniques, Oxford University Press.
- Bowen, W. D., Baker, J. D., Boyd, I. L., Estes, J. A., Ford, J. K. B., Kraus, S. D., Siniff, D., Stirling, I., and Wells, R. S. 2010. Long-term studies, p. 283-305. In Boyd, I. L., Bowen, W. D., and Iverson, S. J. (eds.) Marine Mammal Ecology and Conservation: A handbook of techniques, Oxford University Press, Oxford, U. K.
- Bowen, W. D., Beck, C. A., and Austin, D. 2009. Pinniped ecology, p. 852-860. In Encyclopedia of Marine Mammals. Second Edition. W. F. Perrin, B. Wursig, and H. G. M. Thewissen (eds.) Academic Press
- Bowen, W. D., Hammill, M. O., Koen-Alonso, M., Stenson, G., Swain, D.P., and Trzcinski K. 2009. Proceedings of the National Workshop on the Impacts of Seals on Fish Populations in Eastern Canada (Part 2) CSAS, Proceedings 2009/020
- Bowen, W. D., P. Carter, and M. Hammill. 2011. Estimated Grey Seal Diets near Sable Island Derived from Fecal Samples: 1991 to 2010. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/024: iv+7p.
- Bowen, W.D., C. E. den Heyer, J. I. McMillan and M. Hammill. 2011. Pup production at Scotian Shelf grey seal (Halichoerus grypus) colonies in 2010. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/066
- Boyd, I. L., Bowen, W. D., and Iverson, S. J. 2010. Marine Mammal Ecology and Conservation: A handbook of techniques, Oxford University Press. 450p.
- Breed, G. A., I. D. Jonsen, R. A. Myers, W. D. Bowen, and M. L. Leonard. 2009. Sex-specific, seasonal foraging tactics in adult grey seals (Halichoerus grypus) are revealed by behaviour discriminating state-space models. Ecology 90:3209-3221.
- Breed, G. D., Bowen, W. D. and Leonard, M. L. 2011. Development of foraging strategies with age in a long-lived marine predator. Mar. Ecol. Prog. Ser. 431:267-279.
- Brillant, S., and E.A. Trippel. 2010. Elevations of lobster fishery groundlines in relation to their potential to entangle endangered North Atlantic right whales in the Bay of Fundy, Canada. ICES J. Mar. Sci. 67: 355-364.
- Buckman, A.H., Veldhoen, N., Ellis, G., Ford, J.K.B., Helbing, C.C. and Ross, P.S. 2011. PCB-associated changes in mRNA expression in killer whales (Orcinus orca) from the NE Pacific Ocean. Environmental Science &Technology 45(23):10194-202.
- Burns, J.M., N. Skomp, N. Bishop, K. Lestyk, and M. Hammill. 2010. Development of aerobic and anaerobic metabolism in cardiac and skeletal muscles from harp and hooded seals. The Journal of Experimental Biology 213:740-748
- Calambokidis, J., Barlow, J., Ford, J.K.B., Chandler, T.E., and Douglas, A.B. 2009. Insights into the population structure of blue whales in the eastern North Pacific from recent sightings and photographic identification. Marine Mammal Science 25:816-832
- Chambellant, M. and S.H. Ferguson. 2009. Ageing live ringed seals (Phoca hispida): which 1 tooth to pull? Marine Mammal Science 25: 478-486. DOI: 10.1111/j.1748-7692.2008.00269.x
- Chassot, E., D. Duplisea, M.O. Hammill, A. Caskenette, N. Bousquet, Y. Lambert, and G. Stenson. 2009. The role of predation by harp seals (Pagophilus groenlandicus) in the collapse and non-recovery of northern Gulf of St. Lawrence cod (Gadus morhua). Marine Ecology Progress Series 379: 279-297.
- Christensen, J.R., Letcher, J.R., and Ross, P.S. 2009. Persistent or not persistent? Polychlorinated biphenyls are readily depurated by grizzly bears (Ursus arctos horribilis). Environ.Toxicol.Chem. 28: 2206-2215.
- Clark, D.S., Coopper, T., Doherty, A., Ford, J.K.B., Koops, M.A., Mandrak, N.E., Morris, T.J., and Smedbol, R.K. 2009. Terms and Concepts Used in the SARA Program. Canadian Science Advisory Secretariat, Res. Doc 2009/082. vi + 33 p.
- Crawford, W.R. and J.R. Irvine. 2011. State of physical, biological, and selected fishery resources of Pacific Canadian marine ecosystems in 2010. DFO Can. Sci. Advis. Sec.Res. Doc. 2011/054. x + 163 p.
- Cullon, D.L., Yunker, M.B., Alleyne, C., Dangerfield, N.J., O’Neill, S., Whiticar, M.J., and Ross, P.S. 2009. Persistent organic pollutants in chinook salmon (Oncorhynchus tshawytscha): implications for resident killer whales of British Columbia and adjacent waters. Environ.Toxicol.Chem. 28: 148-161.
- Deecke, V.B., Barrett-Lennard, L.G., Spong, P., and Ford, J.K.B. 2010. The structure of stereotyped calls reflects kinship and social affiliation in resident killer whales (Orcinus orca). Naturwissenschaften 97:513-518.
- Desforges, J.P., Ross, P.S., Loseto, L.L. 2012.Transplacental transfer of polychlorinated biphenyls (PCBs) and polybrominated diphenyl ethers (PBDEs) in arctic beluga whales (Delphinapterus leucas). Environmental Toxicology and Chemistry 31(2):296-300.
- DFO. 2011. Recovery Potential Assessment for Northern Bottlenose Whales (Hyperoodon ampullatus) in Canada. DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2011/031
- DFO. 2011. Impacts of Grey Seals on Fish Populations in Eastern Canada. DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2010/071.Diemer, KM, MJ Conroy, SH Ferguson, DDW Hauser, A Grgicak-Mannion, AT Fisk. 2011. Marine mammal and seabird summer distributions and abundances in the fjords of northeast Cumberland Sound, Baffin Island. Polar Biology 34: 41-48. DOI 10.1007/s00300-010-085
- Doniol-Valcroze, T, Hammill, M. O. and Lesage, V. 2010. Harvest advice for Eastern Hudson Bay beluga (Delphinapterus leucas). DFO Can. Sci. Advis. Sec. Res. Doc. 2010/121. iv + 13 p.
- Doniol-Valcroze, T., Hammill, M.O., Lesage, V. 2011. Information on abundance and harvest of eastern Hudson Bay beluga (Delphinapterus leucas). DFO Can.Sci.Advis.Sec., Res.Doc.2010/121. iv + 13 p.
- Doniol-Valcroze, T., Lesage, V., Giard, J., Michaud, R. 2011. Optimal foraging theory predicts diving and feeding strategies of the largest marine predator. Behavioural Ecology 22(4): 880-888.
- Dufresne, M.M., Frouin, H., Pillet, S., Hammill, M.O., Lesage, V., De Guise, S., and Fournier, M. 2010. Comparative sensitivity of two species of seals to several environmental contaminants using in vitro exposure. Marine Pollution Bulletin 60: 344-349.
- Earle, J. A., M. M. Melia, N. V. Doherty, O. Nielsen, and S. L. Cosby. 2010. Identification of Phocine Distemper Virus in Seals off the East Coast of USA in 2006: Genetic Comparison with Viruses from the 1988 and 2002 Epizootics. Emerging Infectious Diseases Vol. 17 215-220.
- Ferguson, S.H. 2010. MMR Freeman and L Foote (eds): Inuit, polar bears, and sustainable use: local national and international perspectives. Human Ecology 38: 463-465.
- Ferguson, S.H. 2010. Review of aerial survey estimates for ringed seals (Pusa hispida) in western Hudson Bay, 2009 and 2010. DFO Can. Sci. Advis. Sec. Sci. Res. Doc. 2010/xxx.
- Ferguson, S.H., B.G.Young. 2011. Aerial survey estimates of hauled-out ringed seal (Pusa hispida) density in western Hudson Bay, June 2009 and 2010. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/029. iv + 12 p.
- Ferguson, S.H., M.C.S.Kingsley and J.W. Higdon. 2012. Killer whale (Orcinus orca) predation in a multi-prey system. Population Ecology 54(1):31-41.
- Ferguson, SH, L Dueck, LL Loseto, SP Luque. 2010. Bowhead whale (Balaena mysticetus) seasonal selection of sea ice. Marine Ecology Progress Series 411: 285-297.
- Ferreira, E.O., L.L. Loseto, S.H. Ferguson.2011. Assessment of growth-layer groups from ringed seal (Pusa hispida) as biomonitors of inter- and intra-annual Hg, d15N and d13C variation. Can. J. Zool. 89: 774-784.
- Forbes, L.B., L. Measures and A. Gajadhar. 2009. Infectivity of Toxoplasma gondii in northern traditional (country) foods prepared with meat from experimentally infected seals. Journal of Food Protection 72: 1756-1760.
- Ford J.K.B., Rambeau A.L., Abernethy R.M., Boogaards M.D., Nichol L.M., and Spaven L.D. 2009. An Assessment of the Potential for Recovery of Humpback Whales off the Pacific Coast of Canada. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/015. iv + 33 p.
- Ford, J.K.B, G.M. Ellis, P.F. Olesiuk, and K.C. Balcomb. 2010. Linking killer whale survival and prey abundance: food limitation in the oceans’ apex predator? Biology Letters, 6:139-142. Published on-line before print September 15, 2009.
- Ford, J.K.B, Wright, B.M., Ellis, G.M., and Candy, J.R. 2010. Chinook salmon predation by resident killer whales: seasonal and regional selectivity, stock identity of prey, and consumption rates. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/101. iv + 43 p.
- Ford, J.K.B. 2009. Killer whale Orcinus orca. In: Perrin, W.F., Wursig, B., and J.G.M. Thewissen (eds.), The Encyclopedia of Marine Mammals Second Edition. Elsevier, New York, NY. P. 650-657.
- Ford, J.K.B. 2009. Dialects. In: Perrin, W.F., Wursig, B., and J.G.M. Thewissen (eds.), The Encyclopedia of Marine Mammals Second Edition. Elsevier, New York, NY.
- Ford, J.K.B., B. Koot, S. Vagle, N. Hall-Patch, and G. Kamitakahara. 2010. Passive acoustic monitoring of large whales in offshore waters of British Columbia. Can. Tech. Rep. Fish. Aquat. Sci. 2898: v + 30 p.
- Ford, J.K.B., G.M. Ellis, C.O. Matkin, M.H. Wetklo, L.G. Barrett-Lennard, and R.E. Withler. 2011. Shark predation and tooth wear in a population of northeastern Pacific killer whales. Aquatic Biology 11:213-224.
- Ford, J.K.B., R.M. Abernethy, A.V. Phillips, J. Calambokidis, G.M. Ellis and L.M. Nichol. 2010. Distribution and relative abundance of cetaceans in western Canadian waters from ship surveys, 2002-2008. Can. Tech. Rep. Fish. Aquat. Sci. 2913: v + 51 p.
- Frie, A.K., K.-A. Fagerheim, M.O. Hammill, F.O. Kapel, C. Lockyer, G.B. Stenson, A. Rosing-Asvid and V. Svetochev. 2011. Error patterns in age estimation of harp seals (Pagophilus groenlandicus): results from a transatlantic, image-based blind-reading experiment using known-age teeth. ICES J. Mar. Sci. 68:1942-1953.
- Frouin H., Lebeuf M., Hammill M.O., Sjare B., and M.Fournier. 2011. PBDEs in serum and blubber of harbour, grey and harp seals pups from Eastern Canada. Chemosphere 82:663-669.
- Frouin, H, Lebeuf M, Hammill M, Masson S, Fournier M . 2010 Effects of individual polybrominated diphenyl ether (PBDE) congeners on harbour seal immune cells in vitro . Marine Pollution Bulletin 60: 291-298
- Frouin, H., L. Menard, L. Measures, P. Brousseau and M. Fournier. 2010. T lymphocyte-proliferative responses of a grey seal (Halichoerus grypus) exposed to heavy metals and PCBs in vitro. Aquatic Mammals 36: 365-371.
- Frouin, H., Lebeuf, M., Hammill, M., Fournier, M. 2010.Toxic effects of polybrominated diphenyl ethers (PBDEs) on harbour seal (Phoca vitulina) immune cells in vitro. Marine Pollution Bulletin 60 :291–298.
- Frouin, H., Lebeuf, M., Hammill, M., Fournier, M. 2010. Phagocytosis in pup and adult harbour, grey and harp seals. Veterinary Immunology and Immunopathology . 134:160-168.
- Frouin, H., Loseto, L., Stern, G., Haulena, M., and Ross, P.S. 2012. Mercury toxicity in beluga whale lymphocytes: Limited effects of selenium protection. Aquatic Toxicology 109: 185-193.
- Gaden A., S.H. Ferguson, L. Harwood, H. Melling, G.A. Stern. 2009. Mercury trends in ringed seals (Phoca hispida) from the western Canadian Arctic since 1973: associations with sea ice duration. Environmental Science and Technology 43: 3646-3651. DOI: 10.
- Garrett, C. and Ross, P. S. 2010. Recovering resident killer whales: A guide to contaminant sources, mitigation, and regulations in British Columbia. Fisheries and Oceans Canada. Canadian Technical Report of Fisheries and Aquatic Sciences No. 2894. 1-224.
- Gervaise, C., Barazzuti, A., Busson, S., Simard, Y., and Roy, N. 2010. Automatic detection of bioacoustics impulses based on kurtosis under weak signal to noise ratio. Applied Acoustics 71: 1020-1026. DOI: 10.1016/j.apacoust.2010.05.009
- Gervaise, C., Stephan, Y., and Simard, Y. 2010. On the use of time-frequency methods in a passive acoustic monitoring system, in Proceedings of ’2010 IEEE International conference on Acoustics, Speech and Signal Processing’. Dallas, Tx, USA. 14-19 March.
- Givens, G. H., Ryan M. Huebinger, John C. Patton, Lianne D. Postma, Melissa Lindsay, Robert S. Suydam, John C. George, Cole W. Matson, J. W. Bickham. 2010. Population genetics of bowhead whales (Balaena mysticetus) in the western Arctic. Arctic 63: 1-12.
- Gosselin, J.-F., Lesage, V., Hammill, M.O. 2009. Index estimates of abundance for beluga in eastern Hudson Bay, James Bay, and Ungava Bay in summer 2008. CSAS Doc. Res. 2009/006: 25 p. Available at http://www.dfo-mpo.gc.ca/csas
- Gosselin, J-F. Lesage, V. et Hammill, MO. 2009. Abundance indices of beluga in James Bay, eastern Hudson Bay and Ungava Bay in 2008. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/006. iv + 25 p.
- Grant, P.B.C., Johannessen, S.C., Macdonald, R.W., Yunker, M., Sanborn, M., Dangerfield, N., Wright, C., and Ross, P.S. 2011. Environmental fractionation of PCBs and PBDEs during particle transport as recorded by sediments in coastal waters. Environ.Toxicol.Chem. 30: 1522-1532.
- Grebner, D.M., Parks, S.E., Bradley, D.L., Miksis-Olds, J.L., Capone, D.E., and Ford, J.K.B. 2011. Divergence of a stereotyped call in northern resident killer whales. J. Acoustical Society America 129:1067-1072.
- Grist, J., S. Josey, L. Boehme, M. Meredith, F.J.M. Davidson, G. Stenson and M.O.Hammill. 2011. Temperature signature of high latitude Atlantic boundary currents revealed by marine mammal-borne sensor and Argo data. Geophysical Research Letters 38:L15601. 6pp.
- Haarr, M.L., L.D. Charlton, J.M. Terhune and E.A. Trippel. 2009. Harbour porpoise (Phocoena phocoena) presence patterns at an aquaculture cage site in the Bay of Fundy, Canada. Aquat. Mamm. 35: 203-211.
- Hammill, M.O., Bowen, W.D.and B. Sjare. 2010. Status of harbour seals (Phoca vitulina) in Atlantic Canada. In: G.Desportes, A. Bjorge, A. Rosing-Asvid, G. Waring and M. Acquarone (eds.) NAMMCO Scientific Publications, Volume 8:175-190.
- Hammill, M. O. Kingsley, M. C. S. Lesage and Gosselin, J.-F. 2009. Abundance of Eastern Hudson Bay belugas. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/009. iv + 22 p.
- Hammill, M. O., Bowen, W. D., and Blanchard, W. 2009. Timing of grey seal pupping on Hay Island. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/091.Hammill, M.O. 2009. Earless seals (Phocidae). W.F. Perrin, B. Wursig and J.G.M. Thewissen (eds). Pages 342-348, In Encyclopedia of Marine Mammals (2nd ed). Elsevier New York. 1316 pp.
- Hammill, M.O. 2009. Ringed seal (Pusa hispida). W.F. Perrin, B. Wursig and J.G.M. Thewissen (eds). Pages 342-348, In Encyclopedia of Marine Mammals (2nd ed). Elsevier Boston. 1316 pp.
- Hammill, M.O. 2010. Feeding by grey seals in the southern Gulf of St Lawrence. CSAS Res Doc. 2010/130.
- Hammill, M.O. and D.P. Swain 2011. A controlled experiment to test the impact of removals of grey seals on the mortality of southern gulf cod. CSAS Res Doc. 2011/013.
- Hammill, M.O. and G.B. Stenson 2010. Comment on “Towards a precautionary approach to managing Canada’s commercial harp seal hunt” by Leaper et al. ICES J. Mar. Sci. 67:321-322.
- Hammill, M.O. and G.B. Stenson. 2009. A preliminary evaluation of the performance of the Canadian management approach for harp seals using simulation studies. CSAS Res Doc. 2009/093.
- Hammill, M.O. and G.B. Stenson. 2009. Abundance of northwest Atlantic harp seals (1952-2010). CSAS Res. Doc. 2009/114.
- Hammill, M.O. and G.B. Stenson. 2010. Pup production of Northwest Atlantic grey seals in the Gulf of St. Lawrence. Canadian Science Advisory Secretariat Res. Doc. 2010/122.
- Hammill, M.O. and G.B. Stenson. 2011. Estimating abundance of Northwest Atlantic harp seals, examining the impact of density dependence. Canadian Science Advisory Secretariat Res. Doc. 2011/011.
- Hammill, M.O. and M.C.S. Kingsley. 2009. Harvest advice for Eastern Hudson Bay belugas. CSAS Res Doc. 2009/89.
- Hammill, M.O., and Lesage, V. 2009. Seasonal movements and abundance of beluga in northern Quebec (Nunavik) based on weekly sightings information. CSAS Doc. Res. 2009/010: 14 p.
- Hammill, M.O., B. Ferland-Raymond, L.-P. Rivest and G. B. Stenson. 2009. Modelling Northwest harp seal populations: modifying an Excel model to R. CSAS Res. Doc. 2009/108.
- Hammill, M.O., W.D. Bowen and S. Sjare. 2011. Status of harbour seals in Atlantic Canada. NAMMCO Scientific publication . pages 175-189, In G. Desportes, a. Bjorne, A. Rosing-Asvid and G.T. Waring. Harbour seals in the North Atlantic and the Baltic. NAMMCO Scientific publications vol. 8.
- Hammill, M.O., W.D. Bowen and W. Blanchard. 2009. Timing of grey seal pupping on Hay Island. CSAS Res Doc 2009/91.
- Hammill, M.O. and G.B. Stenson. 2011. Modelling grey seal abundance in Canadian waters. 2011/014.
- Handegard, N.O., Demer, D.A., Kloser, R., Lehodey, P., Maury, O., and Simard, Y. 2009. Toward a global ocean ecosystem Mid-trophic Automatic Acoustic Sampler (MAAS). Community white paper. OceanObs’09 Venice, Italy. 8 p.
- Hanna, B., Cott, P.A., Joynt, A.A. and Harwood, L.A. 2012. Managing anthropogenic underwater noise in the Northwest Territories, Canada. In: Popper, A.N.and Hawkins, A. (eds.) The Effects of Noise on Aquatic Life. Advances in Experimental Medicine and Biology, Volume 730. Springer, NewYork. pp 625-627.
- Hanson, M.B., Baird, R.W., Ford J.K.B., Hempelmann-Halos, J., Van Doornik, D.M., Candy, J.R., Emmons, C.K., Schorr, G.S., Gisborne, B., Ayres, K.L., Wasser, S.K., Balcomb, K.C., Balcomb-Bartok, K., Sneva, J.G., and Ford, M.J. 2010. Species and stock identification of prey consumed by endangered southern resident killer whales in their summer range. Endangered Species Research 11:69-82.
- Harris, K.A., Nichol, L.M., and Ross, P.S. 2011. Hydrocarbon concentrations and patterns in free-ranging sea otters (Enhydra lutris) from British Columbia, Canada. Environ. Toxicol. Chem. 30(10):2184-93.
- Harris, K.A., Yunker, M.B., Dangerfield, N.J., and Ross, P.S. 2011. Sediment-associated aliphatic and aromatic hydrocarbons in coastal British Columbia, Canada: concentrations, composition, and associated risks to protected sea otters. Environ.Pollut. 159: 2665-2674.
- Harvey, V. and M.O. Hammill. 2010. Variations on spatial distribution on fish abundance in eastern Scotian shelf over the past four decades. CSAS Res Doc. 2010/132.
- Harvey, V. M.O. Hammill, W.D. Bowen. 2010. Spatial overlap between grey seals and their prey in 4VsW. CSAS Res. Doc. 2010/133
- Harvey, V., M.O. Hammill, D. Swain. 2010. Summer Overlap Between a Central-place Forager And Its Prey in the southern gulf of St. Lawrence CSAS Res Doc 2010/131.
- Harvey, V., M.O. Hammill, D.P. Swain, G.A. Breed, C. Lydersen, and K.M. Kovacs. 2010. Winter foraging by a top predator, the grey seal, in relation to the distribution of prey. CSAS Res. Doc. 2010/124
- Harwood, L. A. 2009. Status of anadromous Arctic charr (Salvelinus alpinus) of the Hornaday River, Northwest Territories, as assessed through community-based sampling of the subsistence fishery, August-September 1990-2007. Can. Ms. Rep. Fish. Aquat. Sci. 2890. vii + 34 p.
- Harwood, L.A. 2012. Marine Mammals. In: C. Burn (ed.) Herschel Island - Qikiqtaryuk: A Natural and Cultural History of Yukon's Arctic Island. Wildlife Management Advisory Council (North Slope), Whitehorse, Yukon.
- Harwood, L. A., Sandstrom, S., and Linn, E. 2009. Status of anadromous Dolly Varden (Salvelinus malma) of the Rat River, Northwest Territories, as assessed through sampling of the subsistence fishery (1995-2007). Can. Ms. Rpt. Fish. Aquatic Sci. 2891.
- Harwood, L.A., Smith, T.G., Auld, J.C. 2012. Fall migration of ringed seals (Phoca hispida) through the Beaufort and Chukchi seas, 2001-02. Arctic 65(1):35-44
- Harwood, L.A., Smith, T.G., Melling, H., Alikamik, J., and Kingsley, M.C.S. In press. Ringed Seals and Sea Ice in Canada’s Western Arctic: Harvest-Based Monitoring 1992–2011. Arctic.
- Harwood, L., Joynt, A.A., Kennedy, D., Pitt, R., and Moore, S. E. 2009. Spatial restrictions and temporal planning as measures to mitigate potential effects of seismic noise on cetaceans: a working example from the Canadian Beaufort Sea, 2007-2008. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/040. iv + 14 p.
- Harwood, L.A., and Joynt, A, A. 2009. Factors influencing the effectiveness of Marine Mammal Observers on seismic vessels, with examples from the Canadian Beaufort Sea. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/048. iv + 9 p.
- Harwood, L.A., Auld, J. C., Joynt, A. A., and Moore, S.E. 2010. Distribution of bowhead whales in the SE Beaufort Sea during late summer, 2007-2009. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/111. v+22 p.
- Harwood, L.A., Gordon, D.C. and Johnson, J. 2012. Fishes. In: C. Burn (ed.) Herschel Island - Qikiqtaryuk: A Natural and Cultural History of Yukon's Arctic Island. Wildlife Management Advisory Council (North Slope), Whitehorse, Yukon.
- Heide-Jørgensen, M.P., K.L. Laidre, Ø. Wiig, L. Postma, L.P. Dueck, and L. Bachmann. 2010. Large scale sexual aggregation of bowhead whales. Endangered Species Research 13: 73-78.
- Higdon J.W., D.D.W. Hauser and S.H. Ferguson. 2012. Killer whales (Orcinus orca) in the Canadian Arctic: Distribution, prey items, group sizes and seasonality. Marine Mammal Science 28(2):E93-E109.
- Higdon, J.W. and S.H. Ferguson. 2009. Loss of Arctic sea ice causing punctuated change in sightings of killer whales (Orcinus orca) over the past century. Ecological Applications 19: 1365-1375.
- Higdon, J.W., and S.H. Ferguson. 2011. Reports of humpback and minke whales in the Hudson Bay region, eastern Canadian Arctic. Northeastern Naturalist 18/3: 370-377.
- Himworth, C.G., M. Haulena, D. M. Lambourn, J.K. Gaydos, J. Huggins, J. Calambokidis, J.K.B. Ford, K. Zaremba, and S. Raverty. 2010. Pathology and epidemiology of phocid herpesvirus-1 infections in wild and rehabilitating harbor seals (Phoca vitulina) in the Northeastern Pacific. J. Wildl. Dis. 46:1046-1051.
- Huggins, F. E., S. A. Raverty, O. Nielsen, N. E. Sharp, J. D. Robertson, N. V. C. Ralston. An XAFS Investigation of Mercury and Selenium in Beluga Whale Tissues. Environmental Bioindicators 4:291-302.
- Hugues P. Benoît HP, Swain DP, Bowen WD, Breed GA, Hammill MO, Harvey V. In press. Can predation by grey seals explain elevated natural mortality in three fish species in the southern Gulf of St. Lawrence? Marine Ecology Progress Series 000:000-000.
- Iverson, S. J., Sparling, C. E., Williams, T., Lang, S.L.C., and Bowen, W. D. 2010. Measurement of individual and population energetics of marine mammals, p. 191-221. In Boyd, I. L., Bowen, W. D., and Iverson, S. J. (eds.) Marine Mammal Ecology and Conservation: A handbook of techniques, Oxford University Press.
- Kelly, T.C., Loseto, L.L., Stewart, R.E.A., Yurkowski, M., and Ferguson, S.H. 2010. Importance of Eating Capelin: Unique Dietary Habits of Hudson Bay Beluga. Pages 53-69 in Ferguson, SH, Loseto LL, Malory, ML (editors). 2010. A little less Arctic: changes to top predators in the world’s largest northern inland sea, Hudson Bay. Springer Publishing Company.
- Kingsley, M.C.S., Richard, P. and Ferguson, S.H. 2012. Stock-dynamic model for the northern Hudson Bay narwhal population based on 1982-2008 aerial surveys. DFO Can. Sci. Advis. Sec. Res. Doc. 2012/020. iv + 20 p.
- Klauda, R. J., M. J. Dadswell, R. A. Cunjak, J. E. Cooper, K. L. Beal, and T. S. Avery, editors. 2009. Challenges for Diadromous Fishes in a Dynamic Global Environment. American Fisheries Society, Symposium 69, Bethesda, Maryland.
- Lachmuth, C.L., Alava, J.J. , Hickie, B.E., Johannessen, S.C., Macdonald, R.W., Ford, J.K.B., Ellis, G.M. Gobas, F.A.P.C, and Ross, P.S. 2010. Ocean disposal in resident killer whale (Orcinus orca) Critical Habitat: science in support of risk management. Fisheries and Oceans Canada. Canadian Science Advisory Secretariat: Research Document No. 2010/116.
- Laidre, K., Heide-Jørgensen, M.P., Stern, H, Richard, P. 2012. Unusual narwhal sea ice entrapments and delayed autumn freezeup trends Polar Biology 35:149-154.
- Lang, S. L. C., D. J. Boness, W. D. Bowen and S. J. Iverson. 2010. Primiparous females do not exhibit reduced maternal care in gray seals (Halichoerus grypus). Mar. Mamm. Sci. DOI: 10.1111/j.1748-7692.2010.00443.x
- Lang, S. L. C., Iverson, S. J. and Bowen, W. D. 2009. Repeatability in lactation performance and the consequences for maternal reproductive success in grey seals. Ecology 90:2513-2523.
- Lang, S. L. C., Iverson, S. J. and Bowen, W. D. 2011. The influence of reproductive experience on milk energy output and lactation performance in the grey seal (Halichoerus grypus). PlosOne 6: 1-11, e19487.
- Larrat, S., S. Lair and L. Measures. 2012. Rakemarks on a harbour porpoise (Phocoena phocoena) calf suggestive of a fatal interaction with an Atlantic white-sided dolphin (Lagenorhynchus acutus). AquaticMammals 38(1):86-91.
- Lawson, J.W. 2009. Perception bias corrections for abundance estimates of cetaceans in Newfoundland waters in during the 2007 T-NASS survey SC/17/AE /14, NAMMCO, Quebec City, 7-9 October 2009.
- Lawson, J.W. 2009. The use of sound propagation models to determine safe distances from a seismic sound energy source, DFO Can. Sci. Advis. Sec. Res. Doc., Ottawa, ON. 2009/060. iv + 15p.
- Lawson, J.W., Gosselin, J.-F., Desportes, G., Acquarone, M., Heide-Jørgensen, M.P., Mikkelsen, B., Pike, D., Víkingsson, G., Zabavnikov, V., and Øien, N. 2009. A note on the distribution of short-beaked common dolphins, Delphinus delphis, observed during the 2007 T-NASS (Trans North Atlantic Sightings Survey), International Whaling Commission, Copenhagen, DK. SC/61/SM35. 4p.
- Lawson. J.W., and Gosselin, J.-F. 2009. Distribution and preliminary abundance estimates for cetaceans seen during Canada’s marine megafauna survey - A component of the 2007 TNASS. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/031. vi + 28 p.
- Lebeuf, M., Trottier, S., Noël, M., Raâch, M. and Measures, L. 2009. A twenty year (1987-2007) trend of PBDEs in beluga from the St. Lawrence Estuary, Canada. Dioxin 2009, Organohalogen Compounds 71: 372-376.
- Lenky C, and B. Sjare. (2011). Changes in seal habitat use of nearshore waters around Newfoundland and Sourthern Labrador: implications for potential predation on salmon. The Open Conservation Biology Journal (5)13-24. DOI 10.2174/1874839201105010013
- Lesage, V, Y Morin, È Rioux, C Pomerleau, SH Ferguson, and É Pelletier. 2010. Stable isotopes and trace metals as indicators of diet and habitat use in cetaceans: predicting errors related to preservation, lipid extraction and normalization. Marine Ecology Progress Series 419:249-265.
- Lesage, V., Baillargeon, D., Turgeon, S., and Doidge, D.W. 2009. Harvest statistics for beluga in Nunavik, 2005-2008. CSAS Doc. Res. 2009/007: 21 p.
- Lestyk, K., L. Folkow, A. Blix , M.O. Hammill, J. Burns. 2009. Development of myoglobin concentration and acid buffering capacity in harp (Pagophilus groenlandicus) and hooded (Cystophora cristata) seals from birth to maturity. J. Comp. Physiol. B. 179:985-996
- Lewis, A., M.O. Hammill, Power, D.W. Doidge and V. Lesage. 2009. Movement and aggregation of Eastern Hudson Bay beluga whales (Delphinapterus leucas): a comparison of patterns found through satellite telemetry and Nunavik traditional ecological knowledge. Arctic 62:13-24.
- Loseto, L., T. Wazny, H. Cleator, B. Ayles, D. Cobb, L. Harwood, C. Michel, O. Nielsen, J. Paulic, L. Postma, P. Ramlal, J. Reist, P. Richard, P.S. Ross, S. Solomon, W. Walkusz, L. Weilgart and B. Williams. 2010. Information in support of indicator selection for monitoring the Tarium Niryutait Marine Protected Area (TNMPA). Fisheries and Oceans Canada Canadian Technical Report of Fisheries and Aquatic Sciences No. 094.
- Loseto, L.L., Stern, G.A., Gemmill, B., Deibel, D, Connelly, T., Prokopowicz, A., Fortier, L., Ferguson, S.H. 2009. Summer diet of beluga whales inferred by fatty acid analysis of the eastern Beaufort Sea food web. Journal of Experimental Marine Biology and Ecology. 374: 12-18. doi:10.1016/j.jembe.2009.03.015
- Loseto, L.L. and Ross, P.S. 2011. Organic contaminants in marine mammals: concepts in exposure, toxicity, and management. In Environmental Contaminants in Biota: Interpreting Tissue Concentrations. Edited by W.N.Beyer and J.P.Meador. CRC Press, Taylor & Francis Group, Boca Raton, FL pp. 349-375.
- Luque, S.P. and S.H. Ferguson. 2009. Ecosystem regime shifts have not affected growth and survivorship of eastern Beaufort Sea belugas. Oecologia 160: 367-378. DOI 10.1007/s00442-009-1300-6
- Luque, S.P. and S.H. Ferguson. 2010. Age structure, growth, mortality, and density of belugas (Delphinapterus leucas) in the Canadian Arctic: responses to environment? Polar Biology 33: 163-178.
- Lynch, M., O. Nielsen, P. J. Duignan, R. Kirckwood, A. Hoskins, and J. P. Y. Arnould. 2011. Serologic Survey For Potential Pathogens and Assessment of Disease Risk in Australian Fur Seals. Journal of Wildlife Diseases 47:555-565.
- Lynch, M., P. J. Duignan, T. Taylor, O. Nielsen, R. Kirckwood, J. Gibbons, and J. P. Y. Arnould. 2011. Epizootiology of Brucella Infection in Australian Fur Seals. Journal of Wildlife Diseases 47:352-363.
- Marcoux, M, M Auger-Méthé, E Chmelnitsky, SH Ferguson, MM Humphries. 2011. Local passive acoustic monitoring of narwhals in the Canadian Arctic: A pilot project. Arctic 64: 307-316.
- Marshall, H. D., K. A. Hart, E. S. Yaskowiak, G. B. Stenson, D. McKinnon and E. A. Perry. 2010. Molecular identification of prey in the stomach contents of harp seals (Pagophilus groenlandicus) using species-specific oligonucleotides. Mol. Ecol. Res. 10:181-189.
- Matthews, CJD, SP Luque, SD Petersen, RD Andrews, SH Ferguson. 2011. Satellite tracking of a killer whale (Orcinus orca) in the eastern Canadian Arctic documents ice avoidance and rapid, long-distance movement into the North Atlantic. Polar Biology DOI 10.1007/s00300-010-0958-x
- Morissette, L. and M.O. Hammill. 2011. A preliminary evaluation of the impacts of grey seal, Halichoerus grypus, predation on the 4T ecosystem and possible effects of their removal on cod (Gadus morhua) recovery. CSAS Res Doc. 2011/016
- Morissette, L., M. Castonguay, C. Savenkoff, D. P. Swain, D. Chabot, H. Bourdages, M. O. Hammill and J. M. Hanson. 2009. Contrasting changes between the northern and southern Gulf of St. Lawrence ecosystems associated with the collapse of groundfish stocks. Econorth Proceedings in DSRII. Deep–Sea Research II 56:2117-2131.
- Mos, L., Cameron, M., Jeffries, S.J., Koop, B.F., and Ross, P.S. 2010. Risk-based analysis of PCB toxicity in harbor seals. Integrated Environmental Assessment and Management 6: 631-640.
- Mosnier, A., Lesage, V., Gosselin, J.-F., Lemieux Lefebvre, S., Hammill, M. O., Doniol-Valcroze, T. 2009. Information relevant to the documentation of habitat use by St. Lawrence beluga (Delphinapterus leucas), and quantification of habitat quality. DFO CSAS Research Document 2009/098.
- Mouy, X., Bahoura, M., and Simard, Y. 2009. Automatic recognition of fin and blue whale calls for real-time monitoring in the St. Lawrence. J. Acoust. Soc. Am. 126(6): 2918-2928.
- MPO, 2011. Ajout d’un brise-lame au quai des pilotes des Escoumins, Québec – effets potentiels sur les mammifères marins. Secrétariat canadien de consultation scientifique du MPO, Réponse des Sciences. 2011/007. 11 pp
- MPO, 2011. Réfection des embarcadères de Tadoussac et Baie-Sainte-Catherine, Québec – effets sur les mammifères marins. Secrétariat canadien de consultation scientifique du MPO, Réponse des Sciences. 2011/009. 13 pp.
- Nichol, L.M. 2009. An assessment of some of the factors affecting observer efficacy during cetacean surveys in Canada’s Pacific Region. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/065. iv + 18 p.
- Nichol, L.M., R. Abernethy, L. Flostrand, T.S. Lee and J.K.B. Ford. 2010. Information relevant for the identification of Critical Habitats of North Pacific Humpback Whales (Megaptera novaeangliae) in British Columbia. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/116. iv + 40p.
- Nichol, L.M., Boogaards, M.D., Abernethy, R. 2009. Recent trends in the abundance and distribution of sea otters (Enhydra lutris) in British Columbia. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/016. iv + 16 p.
- Noël, M., Barrett-Lennard, L.G., Guinet, C., Dangerfield, N., and Ross, P.S. 2009. Persistent Organic Pollutants (POPs) in killer whales (Orcinus orca) from the Crozet Archipelago, southern Indian Ocean. Mar.Environ.Res. 68: 196-202.
- Noël, M., Dangerfield, N., Hourston, R.A.S., Belzer, W., Shaw, P., Yunker, M.B., and Ross, P.S. 2009. Do trans-Pacific air masses deliver PBDEs to coastal British Columbia, Canada? Environ.Pollut. 157: 3404-3412.
- Norman, S.A., S. Raverty, E. Zabek, S. Etheridge, J.K.B. Ford, L.M.N. Hoang, and M. Morshed. 2011. Maternal-fetal transmission of Cryptococcus gattii in harbor porpoise. Emerging Infectious Diseases 17: 304-305.
- Nweeia , M. T., Thackeray, J. F., Eichmiller, F., Richard, P, Leclerc, L-M., Lanham, J. and I. Newton. 2009. A note on isotopic analysis of sectioned tusks of the narwhal (Monodon monoceros) and tusk growth rates. Annals of the Transvaal Museum 45: 138-142.
- Olesiuk, P.F., Lawson, J.W., and Trippel, E.A. 2010. Pathways of effects of noise associated with aquaculture on natural marine ecosystems in Canada, DFO Can. Sci. Advis. Sec. Res. Doc., Ottawa, ON. 2010/025.
- Palacios, G., J. F. X. Wellehan, Jr, S. Raverty, A. V. Bussetti, J. Hui, N. Savji, H. H. Nollens, D. Lambourn, C. Celone, S. Hutchison, C. H. Calisher, O. Nielsen and W. I. Lipkin. 2011. Discovery of an orthoreovirus in the aborted fetus of a Steller sea lion (Eumetopias jubatus) Journal of General Virology, 92, 2558–2565.
- Parsons, K.M., Balcomb, K.C. III, Ford, J.K.B., and Durban, J.W. 2009. The social dynamics of southern resident killer whales and conservation implications for this endangered population. Animal Behaviour 77:963-971
- Paulic, J.E., B. Bartzen, R. Bennett, K. Conlan, L. Harwood, K. Howland, V. Kostylev, L. Loseto, A. Majewski, H. Melling, A. Neimi, J.R. Reist, P. Richard, E. Richardson, S. Solomon, W. Walkusz and B. Williams. 2012. Ecosystem Overview Report for the Darnley Bay Area of Interest (AOI). DFO Can. Sci. Advis. Sec. Res. Doc. 2011/062. vi + 63 p.
- Petersen, S.D., M.H.Hainstock and P.J. Wilson. 2010. Population genetics of Hudson Baymarinemammals: current knowledge and future risks. In: S.H.Ferguson, L.L. Loseto, and M.Mallory (eds.) Global Warming and Arctic Marine Mammals. Springer, New York. pp 237-267
- Petersen, S.D., Tenkula, D., and Ferguson, S.H. 2011. Population Genetic Structure of Narwhal (Monodon monoceros).DFO Can.Sci.Advis.Sec.Res.Doc.2011/021.vi + 20 p.
- Petersen, S.D., Tenkula, D., Ferguson, S.H., Kelley, T., and Yurkowski, D.J. 2012. Sex determination of belugas and narwhals: understanding implications of harvest sex ratio. DFO Can. Sci. Advis. Sec. Res. Doc. 2012/019. iii + 9 p.
- Petersen, S.D., Tenkula, D. and Ferguson, S.H. 2011. Population Genetic Structure of Narwhal (Monodon monoceros). DFO Can. Sci. Advis. Sec. Res. Doc. 2011/021. vi + 20 p.
- Petersen, S.E., S.H.Ferguson.2010. Advice regarding the genetic structure of Canadian narwhal (Monodon monoceros). DFO Can. Sci. Advis. Sec. Sci. Advis. Rep.2011/021.
- Piché, C., L. Measures, C. Bédard and S. Lair. 2010. Bronchoalveolar lavage and pulmonary histopathology in harp seals (Phoca groenlandica) experimentally infected with Otostrongylus circumlitus. Journal of Wildlife Diseases 46: 409-421.
- Pomerleau, C., S.H. Ferguson, W. Walkus. 2011. Stomach contents of bowhead whales (Balaena mysticeus) from four locations in the Canadian Arctic. Polar Biology 34(4):615-620.
- Pomerleau, C., Ferguson, S.H., Luque, S., Patterson, T.A., Lesage, V., Heide-Jørgensen, M.-P., Dueck, L.L. 2011. Bowhead whale Balaena mysticetus diving and movement patterns in the Eastern Canadian Arctic: implications for foraging ecology. Endangered Species Research 15:167-177.
- Pomerleau, C., Winkler, G., Sastri, A., Nelson, J., Vagle, S., Lesage, V., Ferguson, S. 2011. Spatial patterns in zooplankton communities across the eastern Canadian subarctic and Arctic waters: Insights from stable carbon (δ13C) and nitrogen (δ15N) isotope ratios. Journal of Plankton Research. 33(12):1779-1792.
- Prewitt, J.S., Freistroffer, D.V., Schreer, J.F., Hammill, M.O., Burns, J.M. 2010. Postnatal development of muscle biochemistry in nursing harbor seal (Phoca vitulina) pups: Limitations to diving behavior? Journal of Comparative Physiology – B. 180:757–766
- Raach, M., Lebeuf, M. et Pelletier, É. (2011) PBDEs and PCBs in the liver of the St. Lawrence Estuary beluga (Delphinapterus leucas) : A comparison of levels and temporal trends with the blubber. J. Environ. Monit., 13 : 649-656.
- Richard , P.R., Laake, J.L. Hobbs , R.C., Heide-Jørgens en, M.P., Asselin, N.C. and Cleator H. 2010. Baffin Bay Narwhal Population Distribution and Numbers: Aerial Surveys in the Canadian High Arctic, 2002-04. Arctic 63(1): 85-99
- Richard, P. 2009. Marine Mammals of Nunavut. Qikiqtani School Operations, Dept. of Education, Nunavut. 97 p. (2nd reprint with corrections).
- Richard, P.R. 2010. Stock definition of belugas and narwhals in Nunavut. CSAS Research Document - 2010/022: 18 p.
- Richard, P.R. 2010. Survey Index of the Northern Hudson Bay Narwhals, August 2008. CSAS Research Document 2010/021: 21 p.
- Richard, P.R. 2011. Allocation model for landed catches from Baffin Bay narwhal stocks. CSAS Research Document 2011/056: 31 p.
- Ross, P. S. 2010. Impact of at sea disposal on resident killer whale (Orcinus orca) Critical Habitat: Science in support of risk management. Fisheries and Oceans Canada Science Advisory Secretariat: Science Advisory Report No. 046. 1-9 pp.
- Ross, P.S., Couillard, C.M., Ikonomou, M.G., Johannessen, S.C., Lebeuf, M., Macdonald, R.W. and Tomy, G.T. 2009. Large and growing environmental reservoirs of Deca-BDE present an emerging health risk for fish and marine mammals. Marine Pollution Bulletin 58:7-10.
- Ross, P.S., Barlow, J., Jefferson, T.A., Hickie, B.E., Lee, T., MacFarquhar, C., Parsons, E.C.M., Riehl, K., Rose, N., Simpkins, M., Slooten, E., Tsai, J., Wang, J.Y., Wright, A., and Yang, S.-C. 2011. Ten guiding principles for the delineation of priority habitat for coastal cetaceans. Marine Policy 35: 483-488.
- Ross, P.S., Dungan, S.Z., Hung, S.K., Jefferson, T.A., MacFarquhar, C., Perrin, W.F., Riehl, K., Slooten, E., Wang, J.Y., White, B.N., Wursig, B., Yang, S.-C., and Reeves, R.R. 2010. Averting the Baiji syndrome: Characterising habitat for critically endangered dolphins in eastern Taiwan Strait. Aquatic Conservation: Marine and Freshwater Ecosystems 20: 685-694.
- Roux, M.-J., Harwood, L. A., Illasiak, Babaluk, J., and de Graff, N. In press. Fishery resources and habitats in a headwater lake of the Brock River, NT, 2003-2005. Can. Manuscript Report Fisheries and Aquatic Sciences 2932.
- Roy, N., Simard, Y., and Gervaise, C. 2010. 3D tracking of foraging belugas from their clicks: Experiment from a coastal hydrophone array. Applied Acoustics 71: 1050-1056. DOI: 10.1016/j.apacoust.2010.05.008
- Salberg, A-B., T. A. Øigard, G. B. Stenson, T. Haug and K. T. Nilssen. 2009. Estimation of seal pup production from aerial surveys using generalized additive models. Can. J. Fish. Aquat. Sci. 66:847-858.
- Sandstrom, S., Harwood, L. A., and Howland, K. Status of anadromous 2009. Status of anadromous Dolly Varden (Salvelinus malma) of the Rat River, Northwest Territories, as assessed through sampling of the subsistence fishery (1995-2007). Canadian Manuscript Report of Fisheries and Aquatic Sciences 2891.
- Savenkoff, C. Valois, S., Chabot, D. and Hammill, M.O. 2009. Input data and parameter estimates for ecosystem models of the northern Gulf of St. Lawrence. (2003-2005). Can. Tech. Rep. Fish. Aquat. Sci. 2829. 117 p.
- Scheer, J.F., J. L. Lapierre, & M. O. Hammill. 2010. Stomach Temperature Telemetry Reveals that Harbor Seal (Phoca vitulina) Pups Primarily Nurse in the Water. Aquatic Mammals 36:270-277.
- Séquin, G., F. Bouchard, L.N. Measures, C.F. Uhland, and S. Lair. 2011. Infections with Philometra sp. associated with mortalities in wild-hatched captive-raised striped bass, Morone saxatilis (Walbaum). Journal of Fish Pathology 34: 475-481.
- Shelley, L.K., Balfry, S.K., Ross, P.S., and Kennedy, C.J. 2009. Immunotoxicological effects of a sub-chronic exposure to selected current-use pesticides in rainbow trout (Oncorhyncus mykiss). Aquat.Toxicol. 92: 95-103.
- Simard, Y. 2009. Le parc marin du Saguenay–Saint-Laurent: un habitat exceptionnel pour les baleines. Nat. Can. 133(3) : 57-61.
- Simard, Y. 2009. Le Parc Marin Saguenay–Saint-Laurent: processus océanographiques à la base de ce site d’alimentation unique des baleines du Nord-Ouest Atlantique. The Saguenay–St. Lawrence Marine Park: oceanographic process at the basis of this unique forage site of Northwest Atlantic whales. Rev. Sc. Eau / J. Water Sci. 22(2): 177-197.
- Simard, Y. 2009. Passive acoustic monitoring during seismic surveys / Le monitorage par acoustique passive pendant les relevés sismiques. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/068. iv + 22 p.
- Simard, Y. and Harvey, M. 2010. Predation on northern krill (Meganyctiphanes norvegica Sars). In. G.A. Tarling, ed. Biology of Northern Krill. Adv. in Mar. Biol. Vol. 57: 277-306. (Burlington: Academic Press, ISBN: 978-0-12-381308-4)
- Simard, Y., and Sourisseau, M. 2009. Diel changes in acoustic and catch estimates of krill biomass. ICES J. Mar. Sci. 66: 1318-1325. doi: 10.1093/icesjms/fsp055
- Simard, Y., Lepage, R., and Gervaise, C. 2010. Anthropogenic sound exposure of marine mammals from seaways: Estimates for lower St. Lawrence Seaway, eastern Canada. Applied Acoustics 71: 1093-1098. DOI: 10.1016/j.apacoust.2010.05.012
- Simard, Y., Roy, N., Gervaise, C., Giard, S., Conversano M., and Ménard, N. 2010. Passive acoustics applied to white whale density monitoring and 3D tracking their foraging dives in an ecosystem hot spot. Proc. IEEE OES Passive 2010, Brest, France. 23-24
- Simard, Y., Roy, N., Giard, S., Gervaise, C., Conversano, M., and Ménard, N. 2010. Estimating whale density from their whistling activity: example with St. Lawrence beluga. Applied Acoustics 71: 1081-1086. DOI: 10.1016/j.apacoust.2010.05.013
- Sjare B., and Stenson, G.B. 2010 .Changes in the reproductive parameters of female harp seals (Pagophilus groenlandicus) in the Northwest Atlantic. ICES Journal of Marine Science 67:304 -315.
- Spaven, L.D., Ford, J.K.B., and C. Sbrocchi. 2009. Occurrence of leatherback sea turtles off the Pacific coast of Canada, 1931-2009. Can. Tech. Rep. Fish. Aquat. Sci. 2858: 32 p.
- Stenson G. B. 2009. Total removals of Northwest Atlantic harp seals (Pagophilus groenlandicus), 1952-2009. CSAS Res. Doc. 2009/112
- Stenson, G.B., M. O. Hammill and J. Flight.2010. Winter diet of grey seals in Cabot Strait. CSAS Res Doc. 2010/128
- Stenson, G. B., M. O. Hammill and B. Healey. Estimating Reproductive Rates of Northwest Atlantic Harp Seals, 1954-2007, for Population Modelling CSAS Res. Doc. 2009/113
- Stenson, G.B. and M.O. Hammill 2010. Improving the management of Atlantic seals under the precautionary approach. Canadian Science Advisory Secretariat Res. Doc. 2010/135.
- Stenson, G.B. and N. J. Wells. 2011 Current reproductive and maturity rates of Northwest Atlantic harp seals, Pagophilus groenlandicus. Canadian Science Advisory Secretariat Res. Doc. 2010/136.
- Stenson, G.B., M.O. Hammill and B. Healey. 2009. Reproductive rates of Northwest Atlantic harp seals, 1954-2007. CSAS Res. Doc 2009/113.
- Stenson, G.B., Hammill, M.O., and Lawson, J.W. 2009. Estimating pup production of Northwest Atlantic Harp Seals, Pagophilus groenlandicus: Results of the 2008 surveys, DFO Can. Sci. Advis. Sec. Res. Doc., Ottawa, ON. 2009/103. iv + 39p.
- Stenson, G.B., Hammill, M.O., and Lawson, J.W. 2011. How many harp seal pups are there? Additional results from the 2008 surveys 2010/137, DFO Can. Sci. Advis. Sec. Res. Doc., Ottawa, ON. 2010/137. iv + 19p.
- Stenson, G.B., S. Benjamins and D.G. Reddin. 2011 Using bycatch data to understand habitat use of small cetaceans: lessons from an experimental driftnet fishery. ICES J. Mar. Sci. 68:937-946.
- Stevens, T.S., and Lawson, J.W. 2009. Passive acoustic monitoring (PAM) sound catalogue: Approach for NW Atlantic additions, Report to Environmental Studies Research Funds, National Energy Board, Calgary, AB. 28p.
- Stewart, R. E. A. and B. E. Stewart, 2009. Female reproductive systems. Pages 423-428 in Encyclopedia of Marine Mammals, Second Edition. W. F. Perrin, B. Würsig and J. G. M. Thewissen (editors). Academic Press, San Diego. 1316 pp
- Stewart, R.E.A. 2010 Walrus. The Canadian Encyclopedia, The Historica-Dominion Institute, Toronto, http://thecanadianencyclopedia.com April 2010
- Stewart, R.E.A., V. Lesage, J.W. Lawson, H. Cleator and K.A. Martin. 2012. Science Technical Review of the draft Environmental Impact Statement (EIS) for Baffinland’s Mary River Project. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/086. vi + 62 p.Res Doc – submitted Oct 2011
- Stokesbury, M.J.S., Dadswell, M.J., Holland, K.N., Jackson, G.D., Bowen, W.D.and R.K.O’Dor. 2009. Tracking diadromous fishes at sea using hybrid acoustic and archival tags. In: Haro A., Smith K.L., Rulifson R.A., Moffitt C.M., Klauda R.J., et al, (eds). Challenges for Diadromous Fishes in a Dynamic Global Environment. Bethesda: American Fisheries Society Symposium 69.
- Straneo, F., Hamilton, G.S., Sutherland, D.A., Stearns, L.A., Davidson, L., Hammil, M.O., Stenson, G.B. and Rosing-Asvid, A. 2010. Rapid circulation of warm subtropical waters in a major glacial fjord off East Greenland. Nature Geoscience 3:182-186
- Swain, D.P., H.P. Benoit, and M.O. Hammill. 2011. Grey seal reduction scenarios to restore the southern Gulf of St. Lawrence cod population. CSAS Res Doc. 2011/035
- Swain, D. P., Benoît, H. P., Hammill, M. O., McClelland, G., and Aubry, É. 2011. Alternative hypotheses for causes of the elevated natural mortality of cod (Gadus morhua) in the southern Gulf of St. Lawrence: the weight of evidence. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/036. iv + 33 p.
- Templeman, N.D., Campana, S., Colbourne, E., Pepin, P., Power, D., Kulka, D.W., Lawson, J.W., Murphy, E., Simpson, M.R., Miri, C.M., Koen-Alonso, M., Ollerhead, L.M.N., Healey, B., and Wareham, V. 2010. Biophysical Overview of the Laurentian Channel Area of Interest (AOI). DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2010/076.
- Thomas, L., M.O. Hammill and W.D. Bowen. 2011. Assessment of population consequences of harvest strategies for the northwest Atlantic grey seal population. CSAS Res Doc. 2011/007
- Thomas, L., M.O. Hammill and W.D. Bowen. 2011. Estimated size of the Northwest Atlantic grey seal population 1977-2010. CSAS 2011/017
- Tierney, K.B., Baldwin, D.H., Hara, T.J., Ross, P.S., Scholz, N.L., and Kennedy, C.J. 2010. Review: Olfactory toxicity in fishes. Aquat. Toxicol. 96: 2-26.
- Tollit, D., Pierce, G.J., Hobson, K. A., Bowen, W. D., and Iverson, S. J. 2010. Diet, p. 165-190. In Boyd, I. L., Bowen, W. D., and Iverson, S. J. (eds.) Marine Mammal Ecology and Conservation: A handbook of techniques, Oxford University Press.
- Tomy G.T., K. Pleskach, S.H. Ferguson, J. Hare, G. Stern, G. MacInnis, C.H. Marvin and L. Loseto. 2009. Trophodynamics of some PFCs and BFRs in a western Canadian Arctic marine food web. Environmental Science and Technology 43:4076-4081. DOI: 10.1021/es9
- Treble, M.A. and R.E.A. Stewart. 2009. Impacts and risks associated with a Greenland Halibut (Reinhardtius hippoglossoides) gillnet fishery in inshore areas of NAFO Subarea 0. DFO Can. Sci. Advis. Sec. Res. Doc. 2010/032. vi + 18 p.
- Trzcinski, M. K., Mohn, R. and Bowen, W. D. 2009. Estimating the impact of grey seals on the Eastern Scotian Shelf and Western Scotian Shelf cod populations. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/052.
- Trzcinski, M.K., R. Mohn, and W.D. Bowen. 2011. Estimating the Impact of Grey Seals on the Eastern Scotian Shelf Cod Population. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/078: vi + 20p
- Tucker, S., W. D. Bowen, S. J. Iverson and G. B. Stenson. 2009. Intrinsic and extrinsic sources of variation in the diets of harp and hooded seals revealed by fatty acid profiles. Can. J. Zool. 87:139-151.
- Tucker, S., W. D. Bowen, and S. J. Iverson, W. Blanchard, and G. Stenson. 2009. Sources of variation in diets of harp (Pagophilus groenlandicus) and hooded (Cystophora cristata) seals estimated from quantitative fatty acid signature analysis (QFASA). Marine Ecology Progress Series, 384: 287-302.
- Turgeon, J. , P. Duchesne, L.D. Postma, and M.O. Hammill. 2009. Spatiotemporal distribution of beluga stocks (Delphinapterus leucas) in and around Hudson Bay: Genetic mixture analysis based on mtDNA haplotypes – CSAS Res. Doc. 2009/011
- Van Opzeeland, I.C., P.J. Corkeron, D. Risch, G. Stenson, S.M. Van Parijs. 2009. Geographic Variation in vocalizations of pups and mother-pup behavior of harp seals Pagophilus groenlandicus. Aquatic Biology 6: 109-120
- Vazquez-Medina, J.P., N.O. Olguın-Monroy, P.D. Maldonado, A. Santamarıa, M. Ko¨ nigsberg, R. Elsner, M.O. Hammill, J.M. Burns, and T. Zenteno-Savın. 2011. Maturation increases superoxide radical production without increasing oxidative damage in the skeletal muscle of hooded seals (Cystophora cristata). Canadian Journal of Zoology, 89:(3) 206-212, 10.1139/Z10-107
- Ward, E.J., Parsons, K., Holmes, E.E., Balcomb, K.C. III, and Ford, J.K.B. 2009. The role of menopause and reproductive senescence in a long-lived social mammal. Frontiers in Zoology, 6:4.
- Watt, C.A. and Ferguson, S.H. 2011. Stable isotope and fatty acid analyses of samples from entrapped narwhals (Monodon monoceros). DFO Can. Sci. Advis. Sec. Res. Doc. 2010/134. iv + 12 p.
- Westdal, K. Richard, P. and J. Orr. 2009. Migration route and seasonal home range of the Northern Hudson Bay narwhal (Monodon monoceros). In: Ferguson, SH, Loseto, LL, and ML Mallory. A little less Arctic: top predators in the world’s largest northern inland sea, Hudson Bay. Springer Publishing Company.
- Wiig, Ø., M.P. Heide-Jørgensen, C. Lindqvist, K.L. Laidre, L.D. Postma, L.P. Dueck and L. Bachman. 2011. Recaptures of genotyped bowhead whales (Balaena mysticetus) in Eastern Canada and West Greenland. Endangered Species Research 14:235-242.
- Wood, S.A., T.R. Frasier, B.A. McLeod, J.R. Gilbert, B.N. White, W.D. Bowen, M.O. Hammill, G.T. Waring, and S. Brault. 2011. The genetics of recolonization: an analysis of the stock structure of grey seals (Halichoerus grypus) in the northwest Atlantic. Can. J. Zool. 89: 490–497.
- Young, B.G., L.L. Loseto, and S.H. Ferguson. 2010. Diet differences among age classes of Arctic seals: evidence from stable isotope and mercury biomarkers. Polar Biology 33: 153-162
- Young, BG, JW Higdon, SH Ferguson. 2011. Killer whale (Orcinus orca) photo-identification in the eastern Canadian Arctic. Polar Research 30: online 1-11.
- Yunker, M.B., Lachmuth, C.L., Cretney, W.J., Fowler, B.R., Dangerfield, N.J., White, L., and Ross, P.S. 2011. Biota – sediment partitioning of aluminum smelter related PAHs and pulp mill related diterpenes by intertidal clams at Kitimat, British Columbia. Mar. Environ. Res. 72: 105-126.
- Yurkowski, D.J., M. Chambellant and S.H. Ferguson. 2011. Bacular and testicular growth and allometry in the aquatic-mating ringed seal (Pusa hispida): evidence of polygyny? Journal of Mammalogy 92:803-810.
- Date de modification :