
Centre d'expertise sur les mammifères marins : Rapport de recherche scientifique 2006-2008
Table des matières
- Remerciements
- Message du directeur du Centre d'expertise sur les mammifères marins
- 1.0 Aperçu
- 2.0 Dynamique des populations
- 2.1 Abondance
- 2.1.1 SPLASH: Étude internationale sur les rorquals à bosse dans le Pacifique Nord (John Ford)
- 2.1.2 Abondance du béluga de l'ouest de la baie d'Hudson (Pierre Richard)
- 2.1.3 Abondance du béluga de l'estuaire du Saint-Laurent (Jean-François Gosselin)
- 2.1.4 Structure par âge, croissance, mortalité et densité de la population de bélugas dans l'Arctique canadien (Steve Ferguson)
- 2.1.5 Réduction soutenue de la croissance démographique du phoque gris (Don Bowen et Mike Hammill)
- 2.1.6 Moment de la mise bas et abondance des phoques à capuchon de l'Atlantique Nord-Ouest (Garry Stenson et Mike Hammill)
- 2.1.7 Abondance de l'otarie de Steller en Colombie-Britannique (Peter Olesiuk)
- 2.1.8 Abondance des baleines boréales de l'est de l'Arctique (Larry Dueck)
- 2.2 Établissement de la structure de la population à l'aide de la génétique moléculaire
- 2.2.1 Différencier les bélugas de l'est de la baie d'Hudson de ceux de l'ouest de la baie d'Hudson au moyen de marqueurs moléculaires (Lianne Postma)
- 2.2.2 La génétique moléculaire pour l'identification des stocks de narvals au Nunavut (Lianne Postma)
- 2.2.3 Modes de différenciation génétique chez les baleines boréales de l'est de l'Arctique canadien et de l'ouest du Groenland (Lianne Postma)
- 2.3 Reproduction et mortalité
- 2.3.1 Tactique d'accouplement des phoques gris mâles selon leur condition: l'importance de la taille du corps (Don Bowen)
- 2.3.2 Évaluation du dénombrement de nouveau-nés d'otaries de Steller à partir de photographies 35 mm prises à angle oblique (Peter Olesiuk)
- 2.3.3 Cycle biologique et dynamique des populations d'épaulards résidents du nord de la Colombie-Britannique (Peter Olesiuk, Graeme Ellis et John Ford)
- 2.4 Rôle des maladies dans les populations de mammifères marins
- 2.1 Abondance
- 3.0 La place des mammifères marins dans l'écosystème
- 4.0 Répercussions anthropiques sur les mammifères marins
- 5.0 Publications de 2006-2008
2.0 Dynamique des populations
2.1 Abondance
2.1.1 SPLASH: Étude internationale sur les rorquals à bosse dans le Pacifique Nord
John Ford

Figure 1. Lieux d'observation de plus de 6 000 rorquals à bosse au large des côtes de Colombie-Britannique, 1982-2007.
À l'instar de nombreuses populations mondiales de grandes baleines, la population de rorquals à bosse dans le Pacifique Nord a été décimée par la chasse à la baleine pratiquée du début jusqu'à la moitié du XXe siècle. Quelque 28 000 rorquals à bosse ont été tués dans l'est du Pacifique Nord seulement, et à la fin de la chasse commerciale à la baleine en 1965, il restait environ 2 000 baleines, peut-être moins. Au cours des dernières années, le rorqual à bosse a présenté des signes de rétablissement, mais on ne dispose d'aucune estimation fiable de l'abondance de la population ou des tendances pour le Pacifique Nord. L'espèce est toujours considérée comme une espèce menacée en vertu de la Loi sur les espèces en péril du Canada.
Au début des années 2000, un groupe de chercheurs spécialistes des baleines de plusieurs pays côtiers du Pacifique, y compris le Canada, a proposé une étude ambitieuse pour déterminer la situation du rorqual à bosse dans le Pacifique Nord. Appelée SPLASH (acronyme formé des mots anglais « Structure of Populations, Levels of Abundance and Status of Humpbacks », structure des populations, niveaux d'abondace et situation des rorquals à bosse), cette initiative est finalement devenue la plus grande étude sur les baleines jamais entreprise. Elle a fait appel pendant trois ans à plus de 400 chercheurs de 10 pays (Canada, États-Unis, Russie, Japon, Mexique, Philippines, Guatemala, Nicaragua, El Salvador et Costa Rica). Cette étude sur le terrain reposait principalement sur deux approches: la photo-identification des baleines à l'aide de marqueurs naturels sur la pointe de la queue, pour déterminer l'abondance de la population et les liens migratoires entre les aires de reproduction et les aires d'alimentation, et une analyse de l'ADN de tissus cutanés prélevés pour déterminer la structure génétique de la population.
L'étude SPLASH a donné d'excellents résultats. Plus de 8 000 rorquals à bosse ont été catalogués. Par suite des analyses des marquages et des recaptures de cet ensemble de données, la population a été estimée à environ 18 000 baleines dans le Pacifique Nord. Des échantillons de peau ont été prélevés sur plus de 6 000 individus et ceux-ci ont révélé une structure génétique complexe dans les diverses régions du Pacifique Nord.

Nageoires caudales de rorquals à bosse et navire de la Garde côtière canadienne
Photo: John Ford
Les scientifiques du MPO ont joué un rôle de premier plan dans l'étude SPLASH, depuis les premières étapes de la planification jusqu'à l'analyse de milliers de photos d'identification, en passant par la recherche intensive sur le terrain au large des côtes de Colombie-Britannique (figure 1). Comme dans d'autres zones du Pacifique Nord, la population de rorquals à bosse qui se nourrit au large de la côte Pacifique du Canada croît rapidement à un taux annuel estimatif de 4 % en raison du recrutement et d'un taux élevé de survie proche de près de 98 % par an. Selon une estimation réaliste, l'abondance de rorquals à bosse dans les eaux de la Colombie-Britannique serait d'environ 2 000 individus.
Les photographies de rorquals à bosse prises en Colombie-Britannique au cours de l'été et de l'hiver ont été comparées avec celles prises dans des aires de reproduction subtropicales, où ces baleines se rassemblent en hiver. La comparaison a révélé que les rorquals à bosse de la Colombie-Britannique migrent principalement vers deux destinations différentes: vers les eaux au large de la côte ouest du Mexique dans le cas des individus au large de l'île de Vancouver, tandis que ceux présents au large de la côte nord de la Colombie-Britannique migrent principalement vers Hawaï.
SPLASH a permis de mieux comprendre la situation des rorquals à bosse dans le Pacifique Nord ainsi que dans les eaux canadiennes. Elle est venue fort à propos puisque le MPO élabore actuellement une stratégie de rétablissement du rorqual à bosse dans les eaux du Pacifique canadien. Le rorqual à bosse semble bien se rétablir, même s'il n'est pas encore hors de danger. Les risques pesant actuellement sur la survie et le rétablissement de la population de rorquals à bosse sont entre autres les collisions avec des navires, l'enchevêtrement dans les engins de pêche, l'augmentation du bruit sous-marin et la disponibilité limitée des proies. Toutefois, aucune de ces menaces ne semble nuire actuellement au taux de croissance soutenu de la population.
2.1.2 Abondance du béluga de l'ouest de la baie d'Hudson
Pierre Richard

Attroupement de bélugas
Photo: Véronique Lesage
La population de bélugas de l'ouest de la baie d'Hudson passe l'été dans les eaux côtières du Manitoba; les plus grands regroupements se retrouvent dans les estuaires des rivières Seal et Churchill et du fleuve Nelson. On a établi deux estimations du nombre d'animaux à la surface (23 000 et 25 100 individus) à partir des relevés photographiques aériens et des observations visuelles effectués en 1987.
Des relevés aériens ont été effectués dans l'ouest et le sud de la baie d'Hudson en 2004. On a utilisé à la fois des relevés visuels aériens par échantillonnage en ligne et des relevés photographiques pour estimer le nombre de bélugas, avec une correction pour tenir compte du biais de disponibilité (animaux manqués par les observateurs car ils étaient sous l'eau). La nouvelle estimation s'élevait à 57 300 (IC de 95 % – 37 700-87 100). Les estimations antérieures par suite du relevé, corrigées pour tenir compte du biais de disponibilité, ont donné des chiffres similaires, ce qui donne à penser que le nombre d'individus n'a pas connu de changements perceptibles depuis 1987.
2.1.3 Abondance du béluga de l'estuaire du Saint-Laurent
Jean-François Gosselin

Photographie du revelé de bélugas
Photo: Jack Orr
Les relevés visant à évaluer l'abondance de bélugas dans le Saint-Laurent ont utilisé diverses méthodes, entre autres des bateaux, des hélicoptères, des aéronefs, des observations visuelles et des relevés photographiques. Le plan de rétablissement du béluga du Saint-Laurent de 1995 recommandait l'adoption d'une méthode standard pour évaluer l'abondance et améliorer la surveillance de la population. Depuis 1988, sept relevés des bélugas dans l'estuaire du Saint-Laurent ont utilisé une méthode de relevé photographique aérien systématique. Le dernier de ces relevés a été réalisé en 2003 et après avoir appliqué un facteur de correction pour tenir compte du nombre d'animaux sous l'eau pendant le survol de l'aéronef, l'indice d'abondance s'élevait à 1 300 (arrondi à la centaine près). Utilisant les estimations photographiques établies depuis 1988, un modèle démographique donne à penser qu'en dépit des 30 années de protection contre la chasse, le béluga de l'estuaire du Saint-Laurent ne montre aucun signe de rétablissement, la population comptant environ 1 100 individus (arrondis à la centaine près).
On a observé une variabilité importante entre les indices d'abondance depuis 1988, qui, croit-on, est attribuable aux problèmes rencontrés lorsqu'on essaie d'effectuer un relevé d'une petite population ayant une distribution agrégative. En 2005, on s'est appuyé sur 19 relevés visuels par échantillonnage en ligne effectué pour évaluer l'effet de l'altitude sur les estimations d'abondance utilisant cette méthode, ainsi que cinq relevés visuels par échantillonnage en ligne effectués en 2003 pour établir en partie la variabilité associée à la distribution agrégative des bélugas dans le Saint-Laurent et pour comparer les estimations des deux méthodes de relevés. L'indice d'abondance du relevé photographique de 2003 était inférieur, représentant 68 % de l'estimation combinée des cinq relevés visuels par échantillonnage en ligne de cette même année, mais n'était pas différent des estimations combinées des relevés visuels de 2005. Même si les indices d'abondance des deux méthodes n'étaient pas différents, il faudrait procéder à des comparaisons additionnelles pour assurer la calibration de ces deux techniques.
2.1.4 Structure par âge, croissance, mortalité et densité de la population de bélugas dans l'Arctique canadien
Steve Ferguson

Béluga et son baleineau
Photo: W. Klenner
Les écosystèmes marins polaires et subpolaires affichent les plus grandes variations saisonnières du monde et les plus grands extrêmes dans les caractéristiques physiques de l'environnement. Lorsque l'environnement est à ce point déterminant, les stratégies d'évolution biologique sont souvent considérées comme des adaptations à divers degrés de saisonnalité et/ou de prévisibilité des ressources, en général alimentaires. Les contraintes environnementales devraient croître avec la saisonnalité des ressources, notamment alimentaires, et d'un habitat de reproduction adéquat.
Il est difficile de vérifier les prévisions concernant l'influence des facteurs environnementaux et de la densité en raison de l'envergure limitée des comparaisons de la structure par âge et de la croissance entre les populations de bélugas. Une comparaison détaillée s'impose, puisque deux stocks canadiens sont répertoriés parmi les espèces menacées d'extinction, deux sont considérées comme des espèces menacées et deux font partie des espèces préoccupantes en vertu de la Loi sur les espèces en péril.
Dans cette étude, on a comparé la structure par âge, la répartition selon la longueur corporelle, la mortalité et la densité de quatre populations de bélugas de l'Arctique canadien: baie Baffin, baie Cumberland, ouest de la baie d'Hudson et est de la baie d'Hudson. La croissance, mais non la mortalité, montre une corrélation positive significative avec la latitude. La répartition par âge diffère dans sa forme entre les populations, les animaux évoluant aux latitudes les plus septentrionales (est de la mer de Beaufort) étant les plus vieux et ayant les plus grandes longueurs corporelles à l'âge adulte. Même si les différences dans la sélectivité et la pression de la chasse ont peut-être joué un rôle dans les réponses observées, la variation dans la taille corporelle de l'adulte donne à penser que la saisonnalité environnementale exerce peut-être des contraintes plus fortes sur les caractéristiques du cycle biologique des bélugas, qui augmentent à mesure que l'on progresse vers le nord.
2.1.5 Réduction soutenue de la croissance démographique du phoque gris
Don Bowen et Mike Hammill

Colonie de phoques gris sur la plage nord de l'île de Sable
Photo: Don Bowen
L'une des plus longues séries d'estimations de la production de jeunes phoques gris provient d'une étude amorcée en 1962 à l'île de Sable, en Nouvelle-Écosse. Le nombre de jeunes phoques gris dans l'île de Sable a augmenté de manière exponentielle au cours des quarante dernières années. Dans son aire de distribution au Canada, le phoque gris part à la recherche de nourriture principalement sur la Plateforme Scotian et dans le golfe du Saint-Laurent en dehors de la saison de reproduction. Les plus grandes colonies de reproduction se trouvent sur la banquise dans la partie sud du golfe du Saint-Laurent et sur l'île de Sable.
Un relevé aérien utilisant la photographie numérique a été réalisé dans l'île de Sable en janvier 2007 pour estimer la production de jeunes phoques gris. On a dénombré un total de 48 036 nouveau-nés sur les photographies numériques. Après correction pour tenir compte de la proportion de nouveau-nés morts avant le relevé et de ceux nés après le relevé, la nouvelle estimation était de 54 500 individus. Pour le segment de population ne vivant pas sur l'île de Sable, des relevés aériens visuels par bande ont été réalisés pour les concentrations de mise bas sur la glace du golfe du Saint-Laurent et des dénombrements ont été réalisés sur les îles du golfe et le long de la côte est de la Nouvelle-Écosse. Des recherches aériennes ont également été effectuées le long de la côte sud et sud-ouest de Terre-Neuve et de l'île d'Anticosti. Les dénombrements ont été corrigés pour tenir compte de la proportion de mises bas qui a eu lieu après la réalisation du relevé.
Dans la mesure du possible, les dénombrements ont été ajustés en fonction de la proportion de mises bas qui a eu lieu après la réalisation du relevé. Les changements dans la durée de l'étape du pelage propre à l'âge ont donné lieu à un changement de 17 % dans les estimations de production de nouveau-nés. La production totale de nouveau-nés en dehors de l'île de Sable, arrondie au millier près, est estimée à 13 000 animaux. La proportion de jeunes nés sur l'île de Sable a augmenté, passant de moins de 5 % vers le milieu des années 1980 à environ 70 % en 2007.
Selon l'estimation de 2007, la production de nouveau-nés sur l'île de Sable a continué d'augmenter, mais le taux d'accroissement a baissé au cours des deux derniers relevés. La proportion de femelles mettant bas à l'âge de 4 à 7 ans était bien moins grande pour les cohortes de 1998-2002 que pour celle observée de la moitié à la fin des années 1980. La nouvelle estimation de la production de nouveau-nés et les nouvelles données sur l'âge de la première mise bas donnent de nouvelles indications du changement dans les indices vitaux de cette population. La production de nouveau-nés en dehors de l'île de Sable n'a pas affiché la même augmentation que celle observée sur l'île de Sable. Ceci est peut-être dû à des taux de mortalité plus élevés et plus variables des jeunes nés sur la glace dans le golfe du Saint-Laurent.
Les données du relevé ont été intégrées à un modèle démographique en même temps que des données sur les taux de reproduction et sur les prises déclarées. La taille totale estimative de la population à la fin de la saison de reproduction 2007 (c'est-à-dire incluant les nouveau-nés) s'élevait à 304 000 individus.
2.1.6 Moment de la mise bas et abondance des phoques à capuchon dans l'Atlantique Nord-Ouest
Garry Stenson et Mike Hammill

Phoque à capuchon mâle
Photo: Mike Hammill
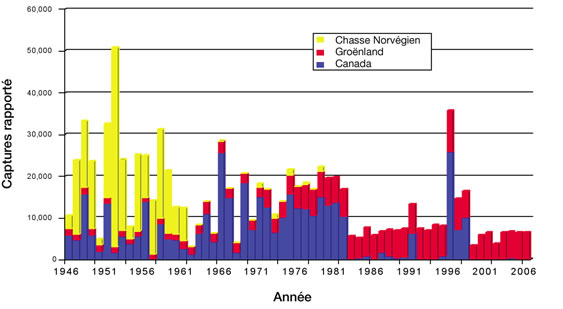
Figure 2. Prises déclarées de phoques à capuchon dans l'Atlantique Nord-Ouest, 1946-2006.
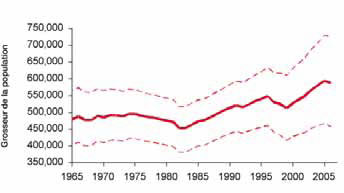
Figure 3. Abondance estimative du phoque à capuchon dans l'Atlantique Nord-Ouest, 1865-2006.
La mise bas des phoques à capuchon a lieu sur la banquise autour de l'île Jan Mayen, dans le détroit de Davis, au large de la côte nord-est de Terre-Neuve (appelée le Front) et dans le golfe du Saint-Laurent. Les relations entre les différents groupes reproducteurs sont mal comprises; certaines analyses utilisant des techniques d'analyse de l'ADN appuient l'hypothèse de l'existence d'une seule population.
Des relevés ont été effectués pendant la saison de reproduction afin d'étudier le moment de la mise bas au printemps. Au Front, la date moyenne de fin de la mise bas était le 28 mars, soit entre le 18 mars et le 4 avril. Dans le golfe, la mise bas prend fin presque tous les ans avant le début d'avril, sauf en 1994, où le modèle a prédit que la mise bas se poursuivrait jusqu'en mai. Si l'on exclut ces données, les naissances auraient pris fin le 28 mars, soit au plus tôt le 25 mars et au plus tard le 31 mars.
On a observé des phoques à capuchon femelles allaitant leurs petits au début mars, ce qui indique que la mise bas de ces phoques s'étale sur une plus longue période que ce que l'on a observé chez les phoques du Groenland qui mettent bas dans la même région. Si l'on tient compte de la possibilité de naissances tardives, en particulier pendant les années de glace dense, l'allaitement devrait être terminé vers le 8-10 avril.
Utilisant des analyses séquentielles des populations ou des indices de survie, les chercheurs ont estimé que la production de jeunes phoques à capuchon au Front était d'environ 30 000 individus au début des années 1980. Toutefois, selon des estimations de la production de jeunes établies grâce à des relevés aériens indépendants effectués au milieu des années 1980, la production de jeunes était d'environ 62 000 pendant les années 1980 et de 83 000 en 1990. Malheureusement, on ne connaît pas bien l'abondance historique des animaux dans le détroit de Davis et dans le golfe. Selon un relevé effectué en 1984, la production estimative de nouveau-nés dans le détroit de Davis serait d'environ 18 600 individus. Relativement peu de jeunes sont nés dans le golfe du Saint-Laurent, les estimations des relevés aériens se chiffrant à 2 000 et à 4 000 animaux en 1990 et 1994 respectivement.
Les estimations les plus récentes de la production de nouveau-nés proviennent des relevés du Front effectués en 2004 et des trois aires de mise bas réalisés en 2005. En 2004, la production de nouveau-nés au Front a été estimée à 123 862. En 2005, elle était estimée à 107 013, tandis qu'on estimait que 6 620 jeunes étaient nés dans le golfe. La production de nouveau-nés dans la concentration de mise bas dans le détroit de Davis était estimée à 3 346. Si l'on combine ces régions, on obtient une production estimative de nouveau-nés pour les trois aires de mise bas de l'Atlantique Nord-Ouest s'élevant à 116 900 individus. La comparaison avec des estimations antérieures donne à penser que la production de jeunes a peut-être augmenté depuis le milieu des années 1980.
Un modèle démographique incorporant les estimations de production des jeunes phoques à capuchon depuis les années 1980, les taux de reproduction et la mortalité causée par l'homme (prises déclarées et individus abattus et perdus) a été utilisé pour estimer l'abondance totale pour la période 1965-2005 (figure 2).
Fitting to pup production estimates from all herds and making assumptions about numbers of hooded seals in the Davis Strait herd for years when this area was not included in the survey program, resulted in an estimated total population of 593,500 in 2006 (Fig. 3). However, there is considerable uncertainty associated with these estimates owing to our lack of understanding of the relationship between the Davis Strait, Front and Gulf pupping areas, few surveys of all three areas, limited reproductive data and uncertain harvest statistics.
2.1.7 Abondance de l'otarie de Steller en Colombie-Britannique
Peter Olesiuk

Abondance de l'otarie de Steller en Colombie-Britannique
Photo: croisière de recherche UBC-MPO
Les otaries de Steller se reproduisent le long du littoral du Pacifique Nord. On connaît deux populations distinctes: une population de l'ouest qui se reproduit en Russie, dans les îles Aléoutiennes et dans le golfe de l'Alaska, et une population de l'est, qui se reproduit dans le sud-est de l'Alaska, en Colombie-Britannique, dans l'Oregon et dans le centre-nord de la Californie. Le MPO effectue des relevés aériens à intervalles de deux à cinq ans depuis le début des années 1970 afin de surveiller les populations de la Colombie-Britannique. Depuis 1994, les relevés du MPO ont été coordonnés avec ceux d'organismes de l'Alaska, de Washington, de l'Oregon, de la Californie et de la Russie afin d'obtenir des estimations sur l'abondance pour l'ensemble de l'aire de répartition de l'espèce.
Les relevés ont lieu entre la fin de juin et le début de juillet, afin de coïncider avec la fin de la saison de mise bas. La plupart des jeunes sont alors nés, mais ils sont encore trop jeunes pour avoir quitté les roqueries, si bien que les relevés fournissent une estimation de la production totale annuelle de nouveau-nés. Des levés des échoueries utilisées par des animaux non reproducteurs sont également effectués. Même si certains individus autres que des jeunes se trouvent en mer et ne sont pas comptabilisés pendant les relevés, ces derniers fournissent de l'information sur la répartition relative des animaux et l'importance des échoueries.
Selon les relevés effectués en Colombie-Britannique, la tendance est à la hausse tant pour ce qui est de la production de jeunes que des dénombrements d'individus autres que des jeunes depuis le début des années 1970, la plupart des augmentations ayant eu lieu depuis les années 1980. L'abondance de l'otarie de Steller a également augmenté dans le sud-est de l'Alaska, l'État de Washington et l'Oregon. Comme les plus grandes roqueries se trouvent près de la frontière entre la Colombie-Britannique et le sud-est de l'Alaska, il est difficile de séparer les populations de ces deux régions, mais toutes deux affichent des tendances à la hausse. En fait, l'abondance combinée des otaries de Steller en Colombie-Britannique et dans le sud-est de l'Alaska a presque quadruplé depuis que l'espèce est protégée, soit depuis le début des années 1970.
Même si des relevés n'ont pas été réalisés à l'échelle de la Colombie-Britannique avant le début des années 1970, il existe des dénombrements pour des roqueries de reproduction datant du début des années 1900. Selon des relevés récents, une proportion relativement constante de la population totale se trouve sur les roqueries, si bien que les dénombrements historiques qui y sont effectués constituent probablement un indice relativement fiable des tendances démographiques globales. Les dénombrements historiques indiquent que l'abondance des otaries en Colombie-Britannique au début des années 1970 a diminué pour s'établir à environ 25-33 % du nombre observé quand les premiers relevés ont été effectués en 1913. Les baisses sont attribuables aux programmes de contrôle des prédateurs et aux pêches expérimentales. Environ 55 000 otaries de Steller ont été tuées en Colombie-Britannique entre 1912 et 1968, ce qui a éliminé une roquerie dans l'archipel Sea Otter Group au large des côtes du centre de la Colombie-Britannique, et réduit les nombres sur les trois roqueries restantes.
Les récentes augmentations en Colombie-Britannique et dans les eaux adjacentes reflètent le rétablissement de populations qui avaient été décimées avant d'être protégées au début des années 1970. L'espèce a recommencé à se reproduire dans l'archipel Sea Otter Group et en 2006, un nombre suffisant de jeunes y sont nés pour qu'on puisse à nouveau la qualifier de roquerie. Ces dernières années, les populations d'otaries de Steller en Colombie-Britannique et dans le sud-est de l'Alaska semblent avoir dépassé les niveaux record historiques et on ne connaît pas la raison de cette augmentation soutenue. En 2002, la population de l'est totale des otaries de Steller était estimée à 46 000-58 000, dont 34 % se reproduisent sur la côte de la Colombie-Britannique.
2.1.8 Abondance des baleines boréales de l'est de l'Arctique
Larry Dueck

Abondance des baleines boréales de l'est de l'Arctique
Photo: Larry Dueck
Il est reconnu que l'abondance des baleines boréales soit difficile à évaluer. Outre leur répartition très étendue et leur densité relativement faible, elles ont aussi tendance à échapper à la vue, sous la surface de l'eau, et sont souvent difficiles à distinguer en raison de leur couleur foncée. Il faut effectuer des relevés dans un délai aussi court que possible, et les conditions atmosphériques et de visibilité compliquent encore la tâche, si bien que des parties importantes du secteur du relevé sont en bout de ligne omis.
Les relevés antérieurs de baleines boréales dans l'est de l'Arctique ne couvraient que des petites parties de leur aire de distribution en raison du coût des relevés aériens et avaient tendance à grandement différer, principalement en raison de facteurs liés aux conditions du relevé et au territoire couvert. Les premiers relevés ont également sous-estimé le nombre de baleines, puisqu'ils ne corrigeaient généralement pas les données pour tenir compte des animaux sous l'eau. L'information sur le comportement de la baleine boréale, en particulier le temps passé sous l'eau, n'était pas encore connue à l'époque.
Une importante opération de relevé a pu être réalisée en 2002-2004 grâce au financement du programme des espèces en péril. Pour obtenir la meilleure information qui soit, on a effectué des relevés aériens à partir d'un aéronef Twin Otter, avec deux observateurs de chaque côté de l'appareil. Cette méthode permet une estimation plus complète du nombre d'animaux puisqu'elle permet aux chercheurs de documenter les espèces de baleines aperçues par l'observateur installé à l'avant, mais non par l'observateur installé à l'arrière, et vice versa. La détermination de la proportion d'animaux non repérés (parce qu'ils se trouvaient sous l'eau) est essentielle pour calculer l'abondance des baleines.
L'analyse de ces données, y compris les ajustements pour tenir compte des animaux passés inaperçus et des animaux sous l'eau, indique que l'abondance des baleines boréales dans l'est de l'Arctique se situe entre 4 800 et 43 000 individus, soit un chiffre estimatif moyen de 14 400 baleines. Même si on ne sait pas précisément où se situe le nombre exact à l'intérieur de cette fourchette, l'estimation nous indique que les nombres révèlent des quantités plus élevées qu'on ne le croyait auparavant et vient corroborer les affirmations des Inuits de nombreuses collectivités selon lesquelles leur nombre a augmenté au cours des dernières décennies.
Note en bas de page: Selon un récent examen du Sous-comité scientifique de la Commission baleinière internationale, l'estimation ci-dessus est peut-être par trop optimiste. Un examen est en cours en vue de répondre à ces préoccupations.
2.2 Établissement de la structure de la population à l'aide de la génétique moléculaire
2.2.1 Différencier les bélugas de l'est de la baie d'Hudson de ceux de l'ouest de la baie d'Hudson au moyen de marqueurs moléculaires
Lianne Postma
L'identification des stocks de bélugas dans la baie d'Hudson et ailleurs dans les eaux canadiennes repose fortement sur l'information génétique. Cette information a été utilisée pour établir une distinction entre les rassemblements estivaux de bélugas appartenant à des lignées maternelles communes. Les analyses préliminaires de ces données corroboraient l'existence d'au moins deux stocks de bélugas dans la baie d'Hudson, généralement appelés stocks de l'est et de l'ouest de la baie d'Hudson. On pense que ces stocks se croisent entre eux dans les aires d'hivernage dans l'est du détroit d'Hudson et dans la baie de Baffin.

Examen des profils d'ADN en laboratoire
Photo: Denise Tenkula, Melissa Lindsay et Robert Bajno
Des échantillons prélevés sur les prises locales ont révélé un mélange d'animaux en migration et d'animaux en estivage. La première étape consiste à relever le nombre de stocks dans la population en isolant les échantillons estivaux. L'étape suivante consiste à examiner les prélèvements des animaux en migration et à effectuer une analyse du stock mélangé pour quantifier les proportions d'animaux des stocks estivaux qui composaient les prises.
Selon les résultats préliminaires de ces analyses, il existe plus de deux stocks de bélugas dans la baie d'Hudson. Tout porte aussi à croire que les échantillons prélevés dans la baie James représentent une population distincte de bélugas qui ne se croisent pas avec les bélugas capturés à d'autres endroits dans la baie d'Hudson et le détroit d'Hudson.
2.2.2 La génétique moléculaire pour l'identification des stocks de narvals au Nunavut
Lianne Postma

Petit groupe de narvals en eau libre
Photo: Larry Dueck
Les résultats des analyses génétiques d'échantillons prélevés sur des narvals dans l'est de l'Arctique canadien entre 1982 et 2001 ont révélé que la différenciation des stocks de narvals dans l'Extrême-Arctique était faible. Il ressort également que le choix des marqueurs aurait dû être suffisant pour mettre au jour les différences entre les stocks.
Dans le cadre des récents travaux concernant le béluga et le morse, on s'est attaché principalement à accroître la quantité d'information génétique utilisée pour examiner les différences entre les stocks et les comportements migratoires. Ces efforts ont accru la robustesse des analyses statistiques et nous ont beaucoup éclairés sur ces espèces.
Au cours des dernières années, les approches analytiques ont grandement changé. Des méthodes plus puissantes récemment mises au point permettent d'établir de manière plus rigoureuse la structure de la population, de quantifier le flux génétique, de fournir de l'information sur l'importance de la migration actuelle par rapport aux associations historiques entre les populations ainsi que de l'information démographique sur l'historique, la croissance et la variabilité de la population. Ces types d'information sont essentiels pour l'établissement de plans de gestion et de plans de rétablissement des stocks en danger de disparition.
Ces méthodes ont toutefois des limites, dont la grande puissance de traitement et la nécessité de disposer de grands ensembles de données. Dans ce projet, le nombre d'échantillons pertinents analysés a été porté à 20 marqueurs microsatellites pour accroître la quantité d'information génétique disponible. L'accent est mis sur l'analyse de 50 échantillons provenant de chacun des quatre lieux – Repulse Bay, Inlet Pond, Fjord Grise et Kikitarjuaq. Les résultats de cette étude sont attendus.
2.2.3 Modes de différenciation génétique chez les baleines boréales de l'est de l'Arctique canadien et de l'ouest du Groenland
Lianne Postma

Chercheurs du MPO marquant des baleines boréales à partir d'un bateau
Photo: DFO
Des échantillons de biopsie ont été prélevés sur des baleines boréales au Nunavut, au Canada, et dans l'ouest du Groenland, et ont été analysés pour 34 loci de microsatellites d'ADN nucléaire. La taille des échantillons était de 8 pour Pelly Bay, 143 pour Igloolik, 16 pour Repulse Bay, 82 pour Pangnirtung et 80 pour Disko Bay.
Les résultats peuvent indiquer la structure des stocks, mais ils pourraient aussi être attribuables à plusieurs facteurs, notamment la consanguinité, le croisement non aléatoire, la sélection naturelle, les conséquences dues à la forte baisse de la population en raison de la chasse commerciale, ou les erreurs de notation des données. Une analyse de la variance moléculaire (AMOVA) a révélé des différences significatives de fréquences d'allèles entre les échantillons d'années différentes à chaque endroit. Ces résultats sont très probablement influencés par la taille des échantillons et la variation dans les sexes et les âges des échantillons de la collection d'une année à l'autre.
La preuve génétique moléculaire des analyses des microsatellites des échantillons de baleines boréales provenant de la baie de Baffin-détroit de Davis et de la baie d'Hudson-bassin Foxe ne semble pas confirmer l'existence de deux populations distinctes. Cette conclusion est corroborée par les résultats des études de localisation par satellite montrant les importants déplacements de baleines boréales entre ces deux régions. Toutefois, les différences génétiques qui ont été relevées et l'information limitée sur la ségrégation en fonction de l'âge et du sexe entre les lieux d'échantillonnage révèlent l'existence d'une certaine structuration des baleines boréales dans leur aire de distribution dont il faudrait peut-être tenir compte dans la gestion de cette population.
2.3 Reproduction et mortalité
2.3.1 Tactique d'accouplement des phoques gris mâles selon leur condition: l'importance de la taille du corps
Don Bowen

Combat de phoques gris mâles sans gagnant
Photo: Don Bowen
Les phoques gris ont une taille dimorphe, les mâles étant 1,5 fois plus grands que les femelles. Chez les espèces dimorphes, on pense souvent que les grands mâles possèdent un avantage pour obtenir une femelle. Cette étude examinait l'importance de la taille du corps et de la composition du corps en tant qu'éléments déterminants de la tactique d'accouplement adoptée par les mâles. Les observations du comportement, ainsi que les mesures de la dépense d'énergie et du succès de reproduction, ont été effectuées pour 42 individus dont on connaissait l'âge pendant les saisons de reproduction de 1997-2001 sur l'île de Sable, au Canada.
Les mâles ayant une masse corporelle élevée arrivaient sur les lieux de reproduction avec plus de réserves lipidiques et d'énergie et dépensaient moins de protéines corporelles que leurs semblables plus petits. Les mâles ayant un pourcentage moins élevé de réserves lipidiques avaient des taux plus élevés d'absorption d'aliments. On observe une corrélation positive entre la quantité d'énergie corporelle à l'arrivée et la durée de la période d'accouplement. Les mâles qui adoptaient pour principale tactique d'accouplement la défense des femelles étaient plus lourds, ils avaient indubitablement plus de tissus adipeux et d'énergie corporelle et ils s'adonnaient plus longtemps à l'accouplement que les mâles qui n'avaient pas adopté cette tactique. Parmi tous les mâles, ceux de taille intermédiaire s'accouplaient avec un plus grand nombre de femelles et obtenaient le plus haut taux de succès. La taille intermédiaire du corps pourrait bien être optimale pendant l'accouplement en raison d'une plus grande agilité durant le combat entre mâles.
2.3.2 Évaluation du dénombrement de nouveau-nés d'otaries de Steller à partir de photographies 35 mm prises à angle oblique
Peter Olesiuk

Les roqueries d'otaries de Steller sont photographiées pendant que l'avion tourne en rond à un angle abrupt et les petits nouveau-nés foncés sont dénombrés à partir de photos haute résolution 35 mm
Photo: Peter Olesiuk
Pour de nombreux pinnipèdes – et l'otarie de Steller ne fait pas exception –, le dénombrement des nouveau-nés constitue la meilleure méthode pour suivre la tendance démographique et estimer l'abondance. La répartition d'autres classes d'âge a tendance à être fort étendue, et une proportion inconnue est en mer et n'est pas comptabilisée pendant les relevés. Par contre, les petits naissent sur les roqueries traditionnelles et sont confinés à terre pendant le premier mois de leur vie. Les recensements de sites de reproduction peuvent donc fournir des estimations de la production totale de nouveau-nés. À partir de là, des modèles démographiques sont utilisés pour extrapoler la taille totale de la population.
Les nouveau-nés d'otaries de Steller sont difficiles à dénombrer à partir de bateaux ou d'avions. Foncés et petits, ils ont tendance à se fondre avec le substrat, et peuvent être dissimulés derrière des rochers, d'autres animaux ou dans des crevasses. Par le passé, on effectuait les dénombrements de nouveau-nés en faisant débarquer des gens sur les roqueries en pleine saison de reproduction, en éloignant les juvéniles et les adultes de la roquerie et en parcourant la roquerie pour compter le nombre de petits présents. Ces dénombrements au sol causent de très fortes perturbations, déplaçant les mâles de leur territoire de reproduction et séparant les petits de leur mère ou les repoussant vers la mer. D'après une étude qui surveillait les changements dans la roquerie avant et après les dénombrements, un tiers de femelles n'ont pas maintenu le contact avec les petits après la perturbation.
En Colombie-Britannique, on a eu recours à des relevés aériens pour surveiller les tendances touchant les otaries de Steller depuis les années 1970. On a dénombré les nouveau-nés à partir de photographies 35 mm prises à angle oblique à partir d'un aéronef qui tournait en rond au-dessus de la roquerie. Si les relevés aériens réduisent la perturbation associée aux dénombrements au sol, on s'interroge néanmoins sur la précision des dénombrements ainsi effectués. Dans les années 1990, les chercheurs du MPO ont fait équipe avec des scientifiques de l'Alaska Department of Fish and Game (ADFG), du National Marine Mammal Laboratory et du Southwest Fisheries Science Center afin de comparer et de calibrer les techniques de dénombrement des petits d'otaries de Steller. Entre 1994 et 1998, le MPO a effectué des relevés à l'île Forrester dans le sud-est de l'Alaska – où naissent plus de nouveau-nés que partout ailleurs – un jour ou deux avant que l'ADFG n'effectue le dénombrement au sol. Si l'on en croit la comparaison, malgré l'existence d'une corrélation étroite entre les relevés aériens et les relevés au sol, invariablement, environ 20 % des petits n'avaient pas été dénombrés par les photographies 35 mm prises à angle oblique.
Dans une étude subséquente, on a constaté que les photographies verticales de format moyen prises à partir d'appareils-photos installés sous l'appareil fournissent des dénombrements statistiquement indifférenciables des dénombrements au sol. En 1998 et en 2002, un relevé des roqueries de la Colombie-Britannique a donc été effectué à l'aide de photographies verticales de format moyen et de photographies 35 mm prises à angle oblique. On observe une corrélation étroite entre les dénombrements utilisant chacune de ces méthodes, mais un petit nombre (4 %) de petits n'ont pas été comptabilisés dans les photographies prises à angle oblique. On croit qu'une proportion plus petite de nouveau-nés étaient dissimulés et n'apparaissaient pas sur les photographies prises à angle oblique des roqueries de la Colombie-Britannique par rapport celles de l'île Forrester du fait que les sites de reproduction de la Colombie-Britannique semblent plus petits ou s'étendre le long des plages, si bien qu'on peut les survoler et les photographier à des angles plus abrupts. Selon ces études, on applique maintenant un petit facteur de correction (1,05) aux dénombrements de jeunes par photographies 35 mm prises à angle oblique en Colombie-Britannique pour tenir compte des animaux dissimulés. Tous les organismes d'Amérique du Nord ont maintenant adopté la photographie aérienne pour le recensement des nouveau-nés d'otaries de Steller, ce qui réduit grandement les perturbations causées par les chercheurs.
2.3.3 Cycle biologique et dynamique des populations d'épaulards résidents du nord de la Colombie-Britannique
Peter Olesiuk, Graeme Ellis and John Ford

Les épaulards résidents demeurent dans leurs groupes natals à vie et se déplacent tous ensemble
Photo: John Ford
Les études sur les populations d'épaulards et les techniques de photo-identification ont été introduites au début des années 1970 par le regretté Michael A. Bigg. Sceptique au départ, Mike a rapidement compris que chaque baleine pourrait être reconnue grâce aux marques naturelles qui lui étaient propres et pris conscience que la photo-identification constituait un puissant outil pour les études démographiques à long terme. Avant son décès prématuré en 1990, il nous a laissé deux documents d'importance capitale qui allaient tracer la voie pour la prochaine génération de chercheurs spécialistes des baleines. Le premier document sur l'organisation sociale et la généalogie montraient que les épaulards résidents des deux sexes demeurent dans leur groupe natal leur vie durant et décrivaient les relations généalogiques entre les individus de chaque groupe matrilinéaire d'épaulards résidents sur la côte de la Colombie-Britannique.
Ce document a ouvert la voie au deuxième document sur le cycle biologique et la biologie des populations d'épaulards résidents. À l'aide de techniques numériques perfectionnées, fort semblables à celles qu'utiliserait un actuaire pour calculer les risques d'assurance, on a établi les paramètres du cycle biologique propres à l'âge d'après les dossiers d'observation de longue date et on les a intégrés aux tables de survie et aux modèles de projection matricielle. Les modèles indiquaient que les épaulards vivaient plus longtemps et se reproduisaient plus lentement que ce que l'on croyait auparavant. Les femelles atteignent généralement la maturité à 14 ans, produisent en moyenne 5 baleineaux sur une durée de vie reproductive de 25 ans, mais peuvent vivre jusqu'à 80 ans. Les mâles atteignent la maturité sexuelle à 13 ans, mais ne sont pas physiquement adultes avant 18 ans, et peuvent vivre jusqu'à 60-70 ans.
On a continué à effectuer des relevés par photo-identification chaque année, et les données sur les naissances et la mortalité des populations d'épaulards résidents couvrent maintenant plus de trente ans. Cette étude de longue haleine nous a offert la possibilité exceptionnelle d'évaluer la façon dont les épaulards s'en sont tirés tant pendant les périodes favorables de croissance démographique que pendant les périodes moins favorables de déclin de la population. Il est intéressant de noter que la situation démographique dépend principalement des changements dans les taux de survie, les taux de reproduction demeurant relativement stables. La stabilité des paramètres de reproduction est peut-être due au potentiel de reproduction très limité des femelles, qui ne produisent qu'en moyenne 5 baleineaux au cours de leur vie. Plus intéressant encore, les animaux des deux sexes et de tous les âges ont affiché des taux de mortalité élevés pendant des périodes moins favorables où les populations déclinaient. Chez la plupart des autres espèces, les jeunes et les animaux inexpérimentés, ou les animaux âgés et frêles sont les plus vulnérables pendant des périodes moins favorables. Cette tendance chez les épaulards peut s'expliquer du fait que, contrairement à d'autres espèces, les animaux des deux sexes et de tous les âges se déplacent ensemble dans leur groupe natal, connaissent les mêmes conditions et souffrent du même manque de proies.
2.4 Rôle des maladies dans les populations de mammifères marins
2.4.1 Maladies importantes pour les populations sauvages de mammifères marins
Lena Measures

Diapositive sur l'histologie de la maladie de Carré du furet
Photo: Lena Measures
Les agents pathogènes les plus importants à l'origine de mortalités massives dans les populations de mammifères marins à travers le monde sont des morbillivirus comme le virus de la maladie de Carré (VMC) infectant les pinnipèdes et le morbillivirus des cétacés (MVC) infectant les cétacés. Le VMC a été responsable de la mort d'environ 20 000 phoques communs et de plusieurs centaines de phoques gris en Europe en 1987-1988 et en 2002. Le MVC a tué des dauphins en Europe et dans l'est des États-Unis, mais le nombre de morts semble plus limité. Le VMC est répandu chez les pinnipèdes canadiens, notamment les phoques du Groenland et les phoques gris, qui seraient les réservoirs du virus. L'absence de mortalité à grande échelle parmi les populations canadiennes de pinnipèdes serait due à l'immunité naturelle du troupeau.
On a signalé quelques cas de mortalité dus au VMC parmi les phoques communs dans la région de la Nouvelle-Angleterre, les plus récents remontant à 2006-2007. On a rapporté la mort d'un seul phoque du Groenland attribuable au VMC dans les eaux canadiennes. Des anticorps contre le VMC avaient été décelés chez les pinnipèdes canadiens dès 1972, mais le virus n'a pas été isolé chez les pinnipèdes canadiens malgré de récents efforts, et aucune étude sur sa pathogénicité ou sa pathogénie n'a été réalisée chez les phoques du Groenland ou les phoques gris. Des études effectuées récemment par Ole Nielsen et Lena Measures du MPO, et des collaborateurs du Centre national des maladies animales exotiques, de l'Agence canadienne d'inspection des aliments et de l'Université de Montréal ont examiné une nouvelle méthode pour isoler et caractériser un isolat de VMC d'une épizootie du phoque survenue en 1988 en Europe en utilisant des furets infectés à des fins expérimentales. Une lignée cellulaire Vero transfectée de manière stable (Vero.DogSLAMtag) exprimant les « molécules canines induisant des signaux d'activation dans les lymphocytes » (SLAM) ou des récepteurs CD150 offrait une réplication virale plus rapide, ce qui a donné lieu à un isolement primaire du virus dans un délai plus court, avec une plus grande production de virus par rapport aux méthodes traditionnelles de culture cellulaire (cellules rénales primaires du phoque et cellules Vero).
Une méthode d'amplification en chaîne par polymérase avec transcription inverse a été mise au point pour déceler une partie du gène du capside nucléique présent uniquement dans le VMC. Cette méthode a été utilisée pour confirmer l'identité du virus qui a par la suite été isolé chez les furets. Les isolats viraux des furets infectés ainsi que les virus isolés au départ chez un dauphin et un marsouin et maintenus dans des cellules Vero se sont aussi reproduits plus vite et ont produit des titres plus élevés de virus lorsqu'ils se sont propagés dans les cellules Vero.DogSLAMtag.
2.4.2 Zoonotic Diseases of Marine Mammals
Lena Measures

Trophozoïtes Giardia
Photo: L. Measures and A. Applebee
Giardia et Cryptosporidium, parasites protozoaires que l'on sait à l'origine des maladies entériques chez les espèces sauvages terrestres, sont également zoonotiques, c'est-à-dire capables d'infecter les humains. De récentes études ont révélé la présence de kystes de Giardia dans les fèces de pinnipèdes canadiens de l'ouest de l'Arctique et de la côte est du Canada. L'objectif de cette nouvelle étude menée à l'Institut Maurice-Lamontagne par Lena Measures et Amber Applebee, dans le cadre de ses recherches doctorales à l'Université Murdoch, à Perth, en Australie, visait à déterminer la prévalence et le génotype de Giardia et Cryptosporidium dans deux espèces de pinnipèdes, le phoque du Groenland (Phoca groenlandica) et le phoque à capuchon (Cystophora cristata), du golfe du Saint-Laurent.
Des échantillons de fèces ont été prélevés sur des phoques adultes et nouveau-nés et examinés en vue de déceler la présence de kystes de Giardia et d'oocystes de Cryptosporidium à l'aide de la microscopie et de la coloration par immunofluorescence. Des tissus du petit intestin des phoques adultes ont été prélevés et examinés en vue de déceler des infections à l'aide de la technique d'amplification en chaîne par polymérase (ACP). On a trouvé des kystes de Giardia dans les fèces de 42 % (16/38) des phoques du Groenland adultes, mais dans aucuns des nouveau-nés de cette espèce (0/20). Même si l'on n'a pas décelé de kystes de Giardia dans les fèces des phoques à capuchon adultes (0/10) à l'aide de la microscopie, 80 % de leurs tissus intestinaux étaient infectés par Giardia d'après la technique d'ACP.
Tant les phoques du Groenland que les phoques à capuchon ont présentés des infections par la souche zoonotique, Giardia duodenalis (assemblage A), déterminée à l'aide d'une technique d'ACP par amorces incluses en deux étapes pour amplifier le gène codant pour la petite sous-unité d'ARNr Giardia. La présence de Cryptosporidium n'a été décelée à l'aide de la microscopie ou de la technique d'ACP dans aucun des tissus intestinaux prélevés des 68 phoques examinés. On a observé des trophozoïtes de Giardia dans les sections histologiques de tissus intestinaux de phoques du Groenland et à capuchon confirmant le dékystement et la prolifération de Giardia chez les phoques.
2.4.3 Échouages et événements de mortalité chez les mammifères marins – Surveillance et enquêtes
Lena Measures

Globicéphale noir vivant échoué
Photo: Lena Measures

Nécropsie d'un béluga
Photo: Lena Measures
Un programme de surveillance et de nécropsie visant la population menacée de bélugas de l'estuaire du Saint-Laurent (ESL) est mené depuis 1982. On signale en moyenne 15 mortalités chaque année, entre trois et douze carcasses étant transportées à la faculté de médecine vétérinaire de l'Université de Montréal pour qu'y soit pratiquée une nécropsie complète en vue de déterminer la cause de la mort et de prélever des tissus pour divers projets de recherche portant sur la santé, le régime alimentaire et les contaminants chimiques présents dans les animaux échoués de cette population. En 2006 et 2007, on a documenté 11 et 17 mortalités, avec 3 et 10 nécropsies réalisées, respectivement.
Des travaux récents effectués avec Michel Lebeuf au MPO ont montré que certains contaminants présents dans les tissus des bélugas de l'ESL diminuaient, tandis que de nouveaux étaient décelés. Entre 1983 et 2007, on a documenté 365 mortalités. Sur les 148 carcasses (les deux sexes et tous les âges confondus) suffisamment fraîches pour la nécropsie entre 1983 et 2002, on a constaté que l'étiologie infectieuse (infections bactériennes et parasitaires) a causé 38 % des décès, la néoplasie ou le cancer terminal 15 %, et que près de 30 % sont morts de causes inconnues. En stratifiant les groupes d'âge, 58 % des baleineaux (moins d'un an) sont morts à la naissance ou juste après la naissance, 81 % des juvéniles (1 à 10 ou 14 ans, selon le sexe) sont morts d'une maladie infectieuse (56 % en raison d'une pneumonie vermineuse), tout comme 32 % des adultes (plus de 10 ou 14 ans selon le sexe). La néoplasie terminale n'a été observée que chez 22 adultes (18 %), dont la moyenne d'âge était de 33 ans (fourchette de 22-59) (GCC/1 an).
On trouvera un aperçu de la population de bélugas de l'ESL intitulé « Quelles sont les causes de mortalité des bélugas du Saint-Laurent? » dans les articles vedettes des Sciences du MPO à l'adresse http://www.dfo-mpo.gc.ca/science/Publications/article/2007/27-07-2007-fra.htm. Malgré la mise en œuvre de diverses mesures de gestion protectrice, l'absence apparente de prédateurs comme les épaulards et l'application de règlements environnementaux sur les produits chimiques toxiques, la population ne s'est pas rétablie. Cet article porte sur l'importance des maladies dans cette population, en particulier les maladies chroniques comme le cancer touchant des animaux qui peuvent vivre plus de 80 ans.
- Date de modification :