
Centre d'expertise sur les mammifères marins : Rapport de recherche scientifique 2009-2011
Table des matières
- Remerciements
- Message du directeur du CEMAM
- 1.0 Dynamique des populations
- 1.1 Croissance des populations de mammifères marins dépendante et indépendante de la densité (Mike Hammill, Don Bowen, Garry Stenson)
- 1.2 Changements dans les taux de reproduction des phoques du Groenland du nord-ouest de l'Atlantique (G.B. Stenson)
- 1.3 Les changements à long terme des indices vitaux des phoques gris sont liés à une réduction du taux de croissance de la population (Don Bowen)
- 1.4 Reproduction des phoques annelés dans la baie d'Hudson (Steve Ferguson, Magaly Chambellant)
- 1.5 Utilisation de la génétique pour acquérir des connaissances sur les groupes de bélugas dans la baie d'Hudson (Mike Hammill, Lianne Postma)
- 2.0 Rôle des mammifères marins dans l'écosystème
- 3.0 Interactions entre les mammifères marins et les humains
- 4.0 Relations avec les conseils de cogestion
- 5.0 Espèces en péril
- 6.0 Publications de 2009-2011
1.0 Dynamique des populations
1.1 Croissance des populations de mammifères marins dépendante et indépendante de la densité (Mike Hammill, Don Bowen, Garry Stenson)
Les effectifs des populations ont naturellement tendance à changer au fil du temps sous l'influence de facteurs comme la concurrence pour les ressources (p. ex., la nourriture), la prédation, l'immigration, l'émigration et la variabilité environnementale. Un problème qui fait souvent l'objet de discussion est le fait de déterminer si ces changements dans les effectifs constituent le résultat de processus qui dépendent ou non de la densité (figure 1). Les facteurs dépendants de la densité ont une incidence sur les effectifs dont l'ampleur dépend de la taille de la population tandis que les facteurs indépendants de la densité ne sont pas liés à la taille de la population. Les processus limitant les effectifs indépendamment de la densité sont souvent causés par des changements environnementaux comme une fonte soudaine des glaces, qui peut avoir des répercussions catastrophiques pour les espèces de phoques qui ont besoin de la glace comme plateforme pour donner naissance à leurs petits et les élever, ce qui mène à une mortalité élevée des jeunes phoques. El Niño constitue un autre exemple, dans le cadre duquel un changement soudain de la température de l'océan a mené à des pénuries alimentaires à grande échelle, entraînant des famines.
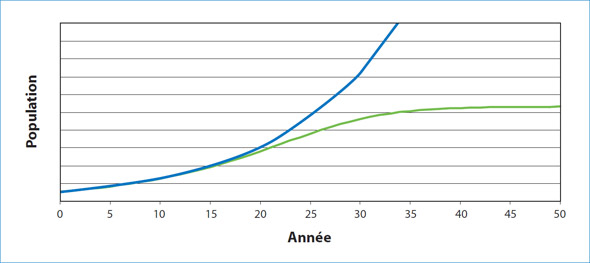
Figure 1. Lorsque la croissance de la population est indépendante de la densité, les ressources ne sont pas limitées et, en théorie, la population augmente de façon continue, souvent de façon exponentielle, jusqu'à ce qu'un événement se produise. Lorsque la croissance de la population est dépendante de la densité, une croissance rapide peut avoir lieu lorsque la population est très petite; toutefois, à un moment donné, la nourriture ou une autre ressource devient limitée et la croissance de la population ralentit et se stabilise à un niveau qu'on appelle la capacité de charge du milieu.
Lorsqu'il y a dépendance à la densité, certaines ressources, habituellement la nourriture (ou l'espace disponible aux fins de reproduction), deviennent de plus en plus limitées; par conséquent, la quantité de nourriture disponible par animal diminue. Sous un certain seuil, cette réduction diminue la condition corporelle, le taux de croissance des individus, le succès reproducteur et en fin de compte la survie. Les effets cette dépendance à la densité ne s'expriment pas uniformément dans la population. Souvent, le premier changement est une augmentation du taux de mortalité des jeunes phoques, surtout au cours de la première année de leur vie, car ils doivent non seulement apprendre à trouver et à capturer de la nourriture, mais doivent aussi consommer plus d'énergie par unité de masse corporelle afin de répondre aux exigences métaboliques et assurer leur croissance. Le taux de survie réduit des jeunes phoques de l'année est souvent associé à une diminution du taux de natalité, car les femelles adultes ont de la difficulté à satisfaire leurs exigences énergétiques de base en plus des coûts supplémentaires de la grossesse et de l'allaitement. Si les conditions deviennent assez graves, la survie des adultes diminue également en réponse au manque de nourriture. Bien que ce modèle général suggère une série de réponses séquentielles, un certain chevauchement aura probablement lieu parmi les changements des indices vitaux pour différentes catégories d'âge et de sexe dans la population.
Lorsque le nombre de naissances dans une population est égal au nombre de décès, la taille de la population se stabilise. Une population stable est considérée comme étant à la capacité de charge du milieu, « K ». Même si l'on tend à la considérer comme étant une capacité constante, K peut varier au fil du temps en réponse à la variabilité environnementale à plus long terme. Par exemple, l'augmentation d'une ressource peut contribuer à améliorer les conditions environnementales, ce qui entraîne une capacité K plus élevée, laquelle a une incidence sur la taille de la population; à l'opposé, des ressources moins élevées peuvent entraîner une diminution de la capacité K.
Il est difficile de détecter lorsqu'une population atteint la capacité de charge ou est en voie de l'atteindre, car certaines fluctuations de la population à court terme sont à prévoir en raison des conditions environnementales variant d'une année à l'autre. De plus, dans le cas des espèces dont nous nous occupons, la taille de la population n'est pas mesurée tous les ans, et chaque estimation de la taille de la population comprend une marge d'erreur. Néanmoins, comme de nombreuses populations de mammifères marins ont été considérablement réduites de façon historique par la chasse, un grand nombre de ces espèces ont augmenté et elles continuent d'augmenter vers la capacité K. Les phoques communs et les baleines grises de la côte ouest du Canada constituent des exemples de populations qui semblent avoir atteint la capacité K ou en être près, tandis que les phoques gris de l'île de Sable présentent des signes de changements dans la croissance de leur population qui sont liés à la densité. Malheureusement, nous n'avons pas de séries à long terme d'estimations d'abondance pour la plupart des populations de mammifères marins; il est donc impossible de déterminer si une population est près de la capacité K ou l'a atteint. De plus, certaines populations sont toujours chassées et il est encore plus difficile de déterminer quelle est la capacité K dans ces situations.
S'il n'y a pas suffisamment de données de relevé pour estimer la capacité K,une autre approche pourrait contribuer à fournir un aperçu de la valeur de la capacité K.
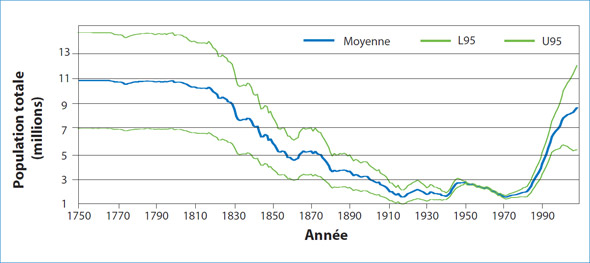
Figure 2: Reconstruction de la population de phoques du Groenland du nord-ouest de l'Atlantique à l'aide de renseignements sur la taille actuelle de la population et de données de chasse datant de la fin des années 1700.
La population de phoques du Groenland du nord-ouest de l'Atlantique a été considérablement réduite en raison de 300 années de chasse. Depuis le début des années 1970, la gestion des quotas a permis à la population d'augmenter et au cours de cette période, nous avons des renseignements sur sa tendance pour une période relativement longue. La population, qui était de moins de deux millions d'animaux au début des années 1970, est passée à près de 8,3 millions d'animaux en 2008; il s'agit de la population la plus importante observée pendant plus de six décennies. Au cours des 30 dernières années, la condition corporelle, la croissance et les taux de reproduction par âge des phoques du Groenland ont montré une tendance au déclin, ce qui suggère que des facteurs dépendants de la densité ont une incidence sur les dynamiques de cette population. Toutefois, des taux de grossesse supérieurs à la moyenne ont entraîné la naissance d'environ 1,6 million de bébés phoques en 2008, ce qui constitue le taux le plus élevé depuis les années 1950. En raison de ces taux de grossesse très variables, il peut être difficile de déterminer si la population est près de sa capacité de charge. Une approche d'estimation de la capacité K serait de tenter de retracer l'histoire de la population jusqu'à une période antérieure au début de la chasse. Les phoques du Groenland font l'objet d'une chasse commerciale depuis le début du 18e siècle. En fait, les chasses aux phoques les plus importantes ont eu lieu au cours de la période des navires de chasse en bois au début des années 1800. Il est très difficile de retracer la population antérieure à la chasse à l'aide des renseignements sur le nombre d'animaux qui ont été pris au fil du temps. Pour conclure que cela reflète la capacité de charge actuelle, il faut supposer que les conditions écologiques qui prévalaient au cours des années 1800 étaient semblables aux conditions environnementales actuelles. Toutefois, lorsque des données de prises historiques ont été inclues dans un modèle et que l'ancienne taille de la population a été retracée, on a pu estimer que la population s'était maintenue à une moyenne de 11 millions d'animaux au début des années 1800. Malheureusement, cette valeur est très incertaine et elle varie possiblement de 7 à 15 millions d'animaux; elle fournit toutefois une indication de la taille maximale possible de cette population (figure 2).
Il est important de comprendre comment les animaux réagissent aux changements de leur abondance. De nombreuses populations de mammifères marins ont été considérablement réduites, et ne comptent aujourd'hui qu'une fraction de leur taille de population vierge. Certaines populations sont maintenant en rétablissement et se rapprochent de leurs abondances historiques. Lorsque les populations sont peu élevées, il y a très peu de différence entre la croissance indépendante de la densité et la croissance dépendante de la densité; dans certains cas, cela a mené à des déclarations affirmant que les populations « explosent » et qu'elles sont hors de contrôle. À un moment donné, les effectifs deviennent toutefois limités par la nourriture, l'espace ou d'autres facteurs, et les facteurs qui dépendent de la densité commencent à limiter la population, ce qui ralentit sa croissance et mène à une stabilisation éventuelle. La compréhension de la nature des changements liés à la mortalité et à la reproduction est essentielle pour l'estimation de l'abondance actuelle et la prévision de la croissance future de la population. La proximité d'une population de la capacité K peut être utilisée en tant qu'indice de sa santé. Une population abondante est plus susceptible de résister aux perturbations soudaines ou aux événements catastrophiques qu'une population plus petite. Les populations saines peuvent également remplir leur rôle écologique traditionnel; pour de nombreux mammifères marins, il s'agit de stabiliser la structure de l'écosystème marin.
1.2 Changements dans les taux de reproduction des phoques du Groenland du nord-ouest de l'Atlantique (G.B. Stenson)
La compréhension des taux de reproduction est essentielle pour comprendre la dynamique des populations de mammifères marins. Comme il est impossible d'étudier la population au complet, nous estimons l'abondance des phoques du Groenland, des phoques à capuchon et des phoques gris en étudiant les petits et en utilisant un modèle de population afin d'extrapoler ces estimations à la taille de la population totale. Par conséquent, la fiabilité de nos estimations de la population dépend de la précision des estimations annuelles du taux de grossesse selon l'âge, d'autant plus que les taux fluctuent en raison de la variation de la taille de l'effectif de la population et des conditions environnementales. Toutefois, le suivi de ces changements est difficile pour la plupart des espèces, car il faut prendre de nombreuses mesures au cours de longues périodes. Depuis les années 1950, des systèmes reproducteurs femelles ont été prélevés sur des phoques du Groenland des eaux de Terre-Neuve et du sud du Labrador; un programme plus systématique entamé dans les années 1980 est toujours en cours aujourd'hui. À l'aide de ces données, les estimations annuelles des taux de grossesse en fin de gestation, de la fécondité (la proportion de femelles adultes qui donnent naissance) et de l'âge moyen de la maturité sexuelle des phoques du Groenland du nord-ouest de l'Atlantique ont été établies.
En général, les taux de grossesse des phoques du Groenland femelles de trois ans étaient très faibles (moins de 10 %); le nombre de femelles enceintes n'est pas élevé. Parmi les phoques de quatre et de cinq ans, les taux de reproduction ont augmenté au cours des années 1970, mais ils ont diminué vers le milieu des années 1980 et sont passés à des niveaux semblables ou inférieurs à ceux des années 1960. Les taux de grossesse pour les femelles de six ans étaient faibles (moins de 67 %) depuis le milieu des années 1990 comparativement aux années précédentes lorsque les taux étaient d'environ 80 %. Parmi les phoques les plus âgés, les taux de grossesse étaient élevés (de 80 à 90 %) jusqu'au milieu des années 1980, mais ils ont ensuite diminué. Depuis, les taux de grossesse ont varié de façon considérable, mais leur moyenne se situe près de 60 %. Depuis 2008, les taux de grossesse ont considérablement diminué; ils sont passés à moins de 30 %. La proportion de femelles adultes qui donnent naissance (le taux de fécondité) suit une tendance semblable; elle est restée relativement élevée jusqu'au milieu des années 1980 puis elle a ensuite diminué (figure 3). Les taux de fécondité récents ont été très variables, les années de faibles taux de grossesse étant associées à des niveaux élevés d'avortements en fin de gestation.

Phoque du Groenland femelle et son petit
Photo : MPO de Terre-Neuve-et-Labrador
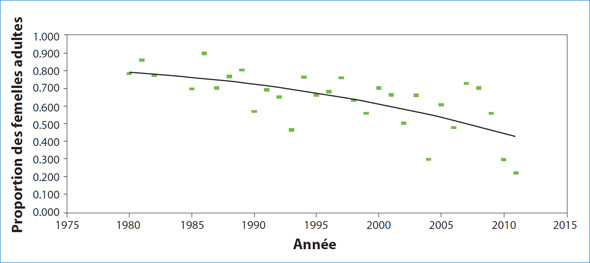
Figure 3: Estimations annuelles de la proportion de phoques du Groenland femelles adultes qui donnent naissance, de 1980 à 2011, en fonction de l'examen des systèmes reproducteurs prélevés au nord-est de Terre-Neuve et au sud du Labrador du mois de décembre au mois de février.

Phoque du Groenland femelle et son petit
Photo : MPO de Terre-Neuve-et-Labrador
L'âge de maturité sexuelle des phoques du Groenland femelles a aussi varié au cours des 50 dernières années. De 1954 à 1976, l'âge moyen de maturité sexuelle était de 5,3 ans. Toutefois, de 1978 à 1987, l'âge moyen de maturité sexuelle des phoques du Groenland femelles est passé à 4,6 ans. En 1990, il a augmenté de près d'une année et au cours de la décennie, l'âge moyen de maturité sexuelle est demeuré relativement stable à 5,6 ans. Sauf en 2000, l'âge moyen de maturité sexuelle a augmenté au début des années 2000 et il a atteint un maximum sans précédent de 6,1 ans en 2005-2006.
Bien que la tendance générale des changements observés corresponde à une réponse de dépendance à la densité de la population, des changements dramatiques dans l'écosystème du nord-ouest de l'Atlantique ont également eu lieu au même moment. Un certain nombre de facteurs physiques et biologiques ont été examinés afin de déterminer ceux qui ont pu avoir une influence sur les taux de grossesse des phoques du Groenland. La diminution générale de la fécondité moyenne semble être un effet de l'augmentation de l'effectif de la population. La variabilité interannuelle est toutefois mieux expliquée par des changements liés aux taux d'avortement en fin de gestation. Les taux d'avortement quant à eux semblent être influencés par les conditions de glace et la disponibilité de la nourriture en début d'hiver, comme l'indiquent les relevés d'abondance de capelans durant les automnes précédents. Cela suggère que la dynamique de population de phoques du Groenland dans le nord-ouest de l'Atlantique est influencée par une interaction complexe de facteurs internes et externes qui comprennent des facteurs physiques et biologiques.
1.3 Les changements à long terme des indices vitaux des phoques gris sont liés à une réduction du taux de croissance de la population (Don Bowen)
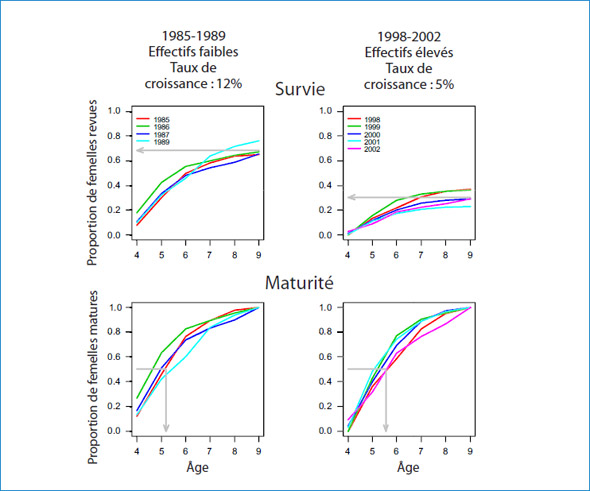
Figure 4: Proportion cumulative des phoques femelles marquées revues sur l'île de Sable entre les âges de 4 et 9 ans (survie, première rangée) et proportion cumulative des femelles revues enceintes ou avec un petit (maturité, deuxième rangée), pour chaque cohorte. Les cohortes marquées de 1985 à 1989, alors que les effectifs étaient faibles et présentaient un taux de croissance de la population élevé, sont situées dans la première colonne, et les cohortes marquées de 1998 à 2002, alors que les effectifs étaient plus élevés et que le taux de croissance était passé de 5 à 4 %, se situent dans la deuxième colonne.
Les populations qui font face à des ressources limitées devraient présenter des changements dans leurs indices vitaux comme une réduction de la survie des juvéniles, un ralentissement de la maturation et une réduction de la survie des adultes. Le taux de croissance de la population des phoques gris (Halichoerus grypus) de l'île de Sable, en Nouvelle-Écosse, a été suivi de 1963 à 2010 en estimant la production de petits. Récemment, le taux d'augmentation de la production de petits est passé de 12 % par année avant 1997 à 4 % par année. De 1969 à 2002, de façon périodique, plus de 7 000 phoques gris ont été marqués d'une manière unique au moment de leur sevrage. Les chercheurs de l'Institut de Bedford (D. Bowen, C. den Heyer et J. McMillan) et de l'Université Dalhousie (S. Iverson) ont recompté ces phoques gris marqués chaque année de 1983 à 2010 en utilisant de trois à cinq recensements hebdomadaires de la colonie de reproduction. Les recaptures de femelles marquées ont été utilisées dans une analyse de capture-recapture afin d'estimer la survie des juvéniles (du sevrage à l'âge de 4 ans) et les probabilités liées à l'âge (de 4 à 13 ans) des primipares. Deux groupes de cohortes (de 1985 à 1989 et de 1998 à 2002) ont été analysés séparément afin de détecter les changements temporels. Les taux estimés de survie des adultes sont toujours demeurés élevés (de 92 à 99 %), mais ils ont légèrement diminué au cours des années 1990. Par contre, les probabilités estimées de la première mise bas aux âges 4 à 13 ans n'ont pas changé, et la survie apparente des juvéniles est passée de 78 % dans les années 1980 à 35 % dans les cohortes les plus récentes (figure 4). Même si le taux de croissance de la population a ralenti de façon considérable, nos résultats indiquent que la survie des adultes est restée élevée. Par conséquent, si les restrictions de nourriture sont responsables du changement observé dans le taux de croissance, la dépendance à la densité est exprimée par la réduction marquée de la survie des juvéniles.
1.4 Reproduction des phoques annelés dans la baie d'Hudson (Steve Ferguson, Magaly Chambellant)
Les phoques annelés figurent parmi les espèces de phoques les plus petites et les plus abondantes, et ont une répartition circumpolaire septentrionale. Les animaux sexuellement matures utilisent principalement la banquise côtière stable avec suffisamment de neige pour établir des abris subnivaux essentiels à la survie des petits. Dans le cadre d'un programme de suivi basé dans les collectivités établi par le MPO de Winnipeg, des échantillons et des mesures de phoques annelés ont été recueillis par les chasseurs Inuits au cours de leur pêche de subsistance d'automne à Arviat, au Nunavut. L'âge des phoques annelés a été déterminé en étudiant les groupes de couches de croissance dans le cément de sections de canines préalablement décalcifiées, coloriées et amincies. Les ovaires et les testicules ont été congelés lorsqu'ils ont été recueillis et ils ont été examinés plus tard aux fins d'évaluation de la reproduction.
La croissance en longueur et en masse (figure 5) des phoques annelés mâles et femelles a été estimée par des courbes de croissance Gompertz. Il n'y avait aucun dimorphisme sexuel portant sur la longueur, la masse ou la condition corporelle (profondeur de la graisse) chez les phoques annelés de l'ouest de la baie d'Hudson. Toutefois, comme les femelles étaient significativement plus âgées, les mâles pourraient être un peu plus grands à un âge donné. Comparativement à d'autres emplacements dans l'Arctique, les phoques annelés de la baie d'Hudson présentaient une longueur et une masse plus petites, ce qui appuie l'hypothèse des différences de taille le long d'un gradient latitudinal.
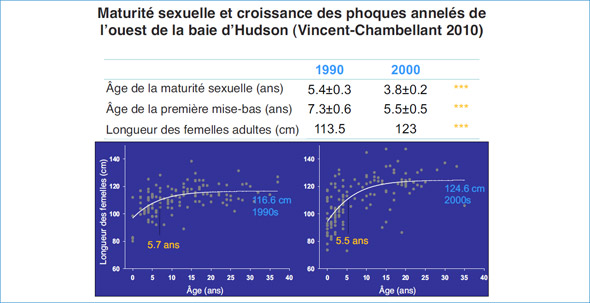
Figure 5: Résumé des changements reproductifs au cours des décennies; les années 1990 par rapport aux années 2000.
Dans la baie d'Hudson, les femelles ont atteint lamaturité sexuelle vers l'âge de six ans,mais lamaturité sexuelle morphologique a pu être détectée à partir de trois ans. Les phoques annelésmâles semblent atteindre lamaturité sexuelle vers cinq ans,mais lamasse des testicules continue d'augmenter jusqu'à l'âge de 10 et 11 ans. Les deux sexes semblent atteindre lamaturité physique avant d'atteindre leurmaturité sexuelle comportementale. Les âges de maturité sont conformes à ceux d'autres phoques annelés dans la baie d'Hudson et dans le reste de l'Arctique,même s'ils se situent dans la partie inférieure du spectre.
Le cycle reproductif des phoques annelés de la baie d'Hudson est semblable à celui des phoques d'autres emplacements de l'Arctique. Les petits naissent sur une banquise côtière ou une banquise stable dans des abris subnivaux qui nécessitent une profondeur de neige de 20 cm ou plus pour fournir une protection suffisante contre la température et les prédateurs de l'Arctique. La date de naissance nominale des petits est établie au 1er avril, mais la période de mise bas peut s'étendre sur plusieurs semaines. Dans la baie d'Hudson, les connaissances traditionnelles et les données récentes convergent vers une période de mise bas antérieure, débutant en février et dont le pic se situe à la mi-mars. Cela appuie l'hypothèse d'un gradient latitudinal des mises bas. Les petits sont sevrés avant la débâcle, après avoir été allaités durant cinq à sept semaines.
On pense que l'accouplement a lieu sous l'eau au moment du sevrage; l'activité sexuelle des mâles atteint un pic signalé de février à avril dans la baie d'Hudson. La gestation des phoques annelés prend environ 10,5 mois, ce qui comprend une période de développement suspendue au cours des premiers deux à trois mois. À la fin du printemps, les phoques annelés entreprennent leur mue annuelle et ils ont besoin d'une banquise pour s'échouer.
La condition corporelle des phoques annelés est la plus médiocre au début de l'été après avoir jeûné pendant les périodes d'accouplement et de mue. Au cours de la période des eaux libres, toutes les catégories d'âges sont mélangées et s'alimentent de façon intensive.
Lorsque la glace commence à se former à la fin de l'automne, les adultes se rassemblent près de la côte afin d'établir leurs territoires. Au cours de cette période, les jeunes sont activement exclus de ces habitats. Les phoques annelés adultes démontrent des signes de fidélité au site au cours de ces mois d'hiver et ils pourraient avoir un système d'accouplement faiblement polygyne avec défense des ressources.
Nous proposons que les dynamiques de la population des phoques annelés suivent un cycle décennal lié aux fluctuations de l'environnement et en particulier au régime de la glace de mer, par l'entremise de changements dans la productivité de l'océan et la pression des prédateurs. Dans les années 2000, les phoques plus jeunes grandissaient plus vite, atteignaient la maturité plus tôt et produisaient plus de petits ayant une meilleure survie que les phoques des années 1990. Des caractéristiques biologiques de ce genre suggèrent davantage une population croissante dans les années 2000 que dans les années 1990. Le changement des caractéristiques biologiques au cours des deux périodes d'étude appuie l'hypothèse que la dynamique de la population des phoques annelés dans l'ouest de la baie d'Hudson peut suivre les variations décennales du régime de glace de mer par l'entremise du forçage atmosphérique (p. ex., oscillation de l'Atlantique Nord). Le cycle de l'effectif des phoques annelés et du rendement reproductif est lié à la sensibilité démographique des petits à la couverture de neige et au besoin de glace stable pour leur survie. Par conséquent, un déclin à long terme de la condition physique des phoques annelés en réponse aux tendances actuelles et prévues des variables environnementales de la baie d'Hudson est probable et il renforce le besoin de données à long terme sur le mode de vie des mammifères marins qui dépendent de la glace et qui vivent à la limite sud de celle-ci.
1.5 Utilisation de la génétique pour acquérir des connaissances sur les groupes de bélugas dans la baie d'Hudson (Mike Hammill, Lianne Postma)
Partout dans leur aire de répartition, les bélugas (Delphinapterus leucas) sont connus pour visiter les estuaires et les embouchures de rivière au cours de l'été. Certains de ces estuaires sont des endroits de regroupements réguliers pour un grand nombre de baleines et ils ont traditionnellement fourni de bonnes occasions pour effectuer une chasse de subsistance. Cela s'explique par le fait que le retour des bélugas est habituellement prévisible et que de nombreuses baleines sont présentes dans un environnement clos qui facilite la chasse. En effet, l'accès fiable à ces sites de regroupement semble avoir été important dans la détermination des patrons de peuplement dans certaines régions de l'Arctique.
On a proposé plusieurs explications pour décrire ce comportement, mais aucune raison ne semble expliquer à elle seule l'existence de ces regroupements. Les estuaires semblent avoir une importance biologique et ils peuvent représenter un habitat essentiel pour ces animaux. Le retour continu des bélugas à ces emplacements et leur persistance à les occuper même s'ils font l'objet d'une perturbation répétée semblent en souligner l'importance.
Les premiers chercheurs à travailler sur ce sujet ont suggéré que ces regroupements de baleines en été étaient formés de groupes distincts d'animaux qui devaient être gérés en tant que populations distinctes ou unités de gestion distinctes. La préoccupation est que si un regroupement est trop chassé, cela peut entraîner l'abandon d'un estuaire. Ce problème peut ne pas avoir de conséquences sur la conservation de l'espèce, mais il peut entraîner une perte de la biodiversité ainsi que des ressources alimentaires pour certains villages situés dans des régions précises.
Afin de déterminer si les bélugas qui vivent dans la baie d'Hudson et dans les alentours sont réellement des groupes distincts de baleines, nous utilisons des marqueurs d'ADN afin de comparer les profils génétiques des animaux. Afin de nous aider, les chasseurs recueillent un échantillon de peau des animaux qu'ils ont chassés et ils nous l'envoient avec des renseignements sur la date et l'emplacement où l'animal a été chassé. Ensuite, nous examinons l'ADN que nous pouvons prélever de l'échantillon de peau et nous cherchons des similitudes et des différences entre les échantillons recueillis de différentes régions au cours de différentes saisons et par différents chasseurs.
Les bélugas sont observés le long de la côte est de la baie d'Hudson, dans la baie James et le long de la côte ouest de la baie d'Hudson au cours des mois d'été. Le groupe de l'est de la baie d'Hudson est le plus petit et il est formé d'environ 3 000 animaux. Les bélugas qui passent l'été à l'ouest de la baie d'Hudson se chiffrent à 57 000, et ils forment peut-être le regroupement de baleines le plus important au monde. Un total de 9 000 autres baleines se trouvent dans la baie James. Les bélugas de la baie d'Hudson migrent à la fin de l'été ou à l'automne vers le détroit d'Hudson, où ils passent l'hiver, même si certains animaux de la baie James semblent rester dans la région de la baie James tout au long de l'année. Les patrons de migration de ces baleines sont très intéressants, car il s'agit du moment où les différentes populations semblent se mélanger et cela leur donnerait l'occasion de s'accoupler. On pense que l'accouplement entre les bélugas a lieu au cours des mois de la fin de l'hiver et du début du printemps. La gestation dure de 9 à 12 mois; les baleineaux naissent le printemps suivant. Les baleineaux restent auprès de leur mère de 12 à 18 mois. Ces caractéristiques biologiques des bélugas contribuent aux types de patrons que nous pouvons constater lorsque nous examinons l'ADN des échantillons fournis par les chasseurs.

Échantillon de Muktuk
Photo: Lisa Loseto
Au cours des analyses génétiques, nous examinons deux types d'ADN : l'ADN mitochondrial et l'ADN nucléaire. L'ADN mitochondrial (ADN mt) est un type d'ADN qui est principalement transmis de la mère à son petit. L'examen des renseignements tirés de l'ADN mt est utile pour l'identification de groupes d'animaux en fonction de leur lignée maternelle. Par contre, l'ADN nucléaire (ADN n) est le type d'ADN qui est le plus fréquemment utilisé dans la science judiciaire, car il est hérité de façon égale de chacun des parents. Pour les bélugas, l'ADN n peut donc être utile pour découvrir des renseignements sur les patrons de reproduction, c.-à-d. les groupes qui s'accouplent ensemble et le type de relations que les baleines individuelles ont entre elles.
Parmi les échantillons d'ADN recueillis des bélugas des côtes ouest et est de la baie d'Hudson et des animaux passant l'hiver dans le détroit d'Hudson, il n'y a que peu de différenciation ou aucune différenciation entre les échantillons à l'aide de l'ADN nucléaire. Cela indique que les animaux qui se reproduisent entre eux le font probablement dans les régions d'hivernage où les animaux ont l'occasion d'être ensemble. Toutefois, les échantillons des animaux provenant des différentes zones d'estivage contenaient des types d'ADN mitochondrial distincts. Les différences d'ADN mt les plus importantes ont révélé que les bélugas capturés en été le long de l'arc de la côte de l'est de la baie d'Hudson (EBH) formaient un groupe génétiquement distinct d'animaux. Sur le plan génétique, ces baleines de l'EBH sont plus liées aux bélugas du fleuve Saint-Laurent et elles sont très différentes des deux principaux groupes de bélugas à proximité, c.-à-d. les baleines de l'ouest de la baie d'Hudson et du sud-est de l'île de Baffin.
À l'aide des signatures génétiques d'ADN mt, on a déterminé que les échantillons de la partie du Nord du Québec (Nunavik) du détroit d'Hudson contiennent une proportion importante (de 7 à 31 %) de bélugas du groupe de l'EBH. Cette proportion est beaucoup plus élevée que les prévisions basées sur la taille relative du groupe de baleines de l'ouest de la baie d'Hudson (57 000 animaux) comparativement au groupe de bélugas de l'est de la baie d'Hudson (3 000 animaux). Cela indique des différences dans les patrons de migration entre les deux groupes, en particulier lorsqu'ils arrivent dans les eaux du détroit d'Hudson.

Extraction d'ADN de la peau
Photo: Lianne Postma
Grâce à un travail collaboratif avec des collègues de l'Université Laval, des analyses plus approfondies de l'ADN nucléaire des échantillons de peau indiquent que les parents et les petits, ainsi que la demi-fratrie et d'autres baleines parentes, se déplacent en restant en association étroite dans l'espace et dans le temps. Des relations plus solides sont signalées entre les mères et leurs filles qu'entre les mères et leurs fils. Ces résultats suggèrent un réseau d'individus liés, en particulier parmi les femelles. Ces réseaux se déplacent entre leurs régions d'été et d'hiver. Par conséquent, les jeunes bélugas sont capables d'apprendre la route migratoire précise de leur mère et de leurs parents, car ils auront effectué de deux à quatre migrations au moment de leur sevrage. Cet apprentissage de routes migratoires précises peut avoir des implications importantes en matière de conservation et de gestion. Si les animaux ont tendance à maintenir les groupes partageant des liens de parenté et à visiter les mêmes régions de façon répétitive, la perte de ces groupes entraînera la perte des connaissances au sujet des routes migratoires. Donc, si les animaux sont éliminés d'un endroit en particulier, les chances de recolonisation de cette zone sont minimes. Cela semble être le cas dans certaines régions du Nord du Québec.
Ces aperçus fournis par la génétique sont très intéressants, et ils sont également soutenus par des renseignements tirés d'autres types d'études comme les observations locales, la télémétrie satellitaire, les analyses des isotopes stables et les comparaisons des signatures de contaminants. Ensemble, les données contribuent à établir une base de connaissances sur les bélugas qui peut être utilisée afin de prédire leur vulnérabilité à divers facteurs de stress dans leur environnement. Conséquemment, les décisions en matière de gestion peuvent être orientées afin de promouvoir la conservation de ces animaux.
- Date de modification :